
秸秆还田下土壤有机质激发效应研究进展
![]()
![]() 第 58 卷 第 6 期 土 壤 学 报 Vol. 58,No. 6
第 58 卷 第 6 期 土 壤 学 报 Vol. 58,No. 6
2021 年 11 月 ACTA PEDOLOGICA SINICA Nov.,2021
![]()
DOI :10.11766/trxb202006260259
张叶叶,莫非,韩娟,温晓霞,廖允成 . 秸秆还田下土壤有机质激发效应研究进展[J]. 土壤学报, 2021 ,58 ( 6 ): 1381– 1392. ZHANG Yeye ,MO Fei ,HAN Juan ,WEN Xiaoxia ,LIAO Yuncheng. Research Progress on the Native Soil Carbon Priming after Straw Addition[J]. Acta Pedologica Sinica ,2021 ,58 ( 6 ): 1381– 1392.
秸秆还田下土壤有机质激发效应研究进展*
张叶叶1 ,莫 非1† ,韩 娟1 ,温晓霞 1 ,廖允成 2†
( 1.西北农林科技大学农学院,陕西杨陵, 712100 ;2. 山西农业大学农学院,山西太谷, 030801 )
摘 要:土壤有机质是农田肥力的基础与核心, 对作物产量、农业环境, 甚至地球碳循环意义重大。作物秸秆作为农田土壤 有机碳库的重要外部补充, 其还田过程对土壤有机碳周转和碳库平衡具有显著影响。激发效应是一种因新鲜有机质输入而导 致土壤本底有机质矿化速率发生改变的现象。秸秆还田导致的土壤有机质分解激发, 不仅涉及秸秆资源化高效利用, 还直接 关系到农田土壤碳库的平衡及其功能, 因此备受科学界关注。尽管对外源有机质输入引起的土壤有机质激发效应的理论研究 已取得了较大进展, 但如何结合最新的理论结果到秸秆还田固碳减排的生产实践中仍面临着较大的挑战, 这主要归结于对农 田土壤有机质分解激发效应的发生特点和规律, 及其背后的土壤、气候、管理等相关的驱动因子和过程还未完全明确。据此, 本文首先对土壤有机质分解激发效应发生的理论研究进展(包括: 共代谢理论、氮矿化理论、化学计量比和微生物残体再利 用)进行了系统综述。其次, 结合已有的研究证据和理论假设进一步概述了秸秆还田过程中影响激发强度和方向的潜在驱动 因素, 如: 秸秆类型和数量、还田方式、水肥管理、土壤属性、气候因子等。最后,从秸秆还田的高效性、农田碳库的可持 续和农业环境的友好性出发, 对秸秆还田土壤有机质分解激发的潜在研究方向进行了展望, 并就秸秆还田改善土壤碳库的优 化措施提出了建议。
关键词: 秸秆还田;激发效应;土壤有机碳
中图分类号: S15 文献标志码: A
Research Progress on the Native Soil Carbon Priming after Straw Addition
ZHANG Yeye1 , MO Fei1† , HAN Juan1 , WEN Xiaoxia1 , LIAO Yuncheng2†
(1 . College of Agronomy, Northwest A&F University, Yangling, Shaanxi 712100, China; 2. College of Agronomy, Shanxi Agricultural University, Taigu, Shanxi 030801, China)
Abstract: Soil organic carbon is the foundation and core of farmland fertility, which is of great significance for crop yield, agricultural environment and the earth's carbon cycling. Crop straw is one of the important external sources for soil organic carbon replenishment in cropping soils. The processes of returning straw to soil exert a significant effect on the turnover of soil organic carbon and hence carbon pool balance. Soil carbon priming is a common phenomenon, which can be defined as an distinct shift of native organic carbon mineralization due to fresh organic matter inputs into the soils. There is an increased attention to the straw-induced priming effect in the recent years, due mainly to the fact that straw incorporation not only involves
* 国家自然科学基金项目(3180101087)资助 Supported by the National Natural Science Foundation of China ( No. 3180101087 ) † 通讯作者 Corresponding author ,E-mail :yunchengliao@163.com ;mofei@nwafu.edu.cn
作者简介:张叶叶( 1995— ) ,女,陕西咸阳人,博士研究生,主要从事农业生态学研究。 E-mail :zhangyeye8296@nwafu.edu.cn
![]() 收稿 日期:
收稿 日期:![]() 2020–06–26 ;收到修改稿 日期:
2020–06–26 ;收到修改稿 日期:![]() 202 1–03–07 ; 网络首发 日期 ( www.cnki.net ) :202 1–03–3 1
202 1–03–07 ; 网络首发 日期 ( www.cnki.net ) :202 1–03–3 1 ![]()
![]()
![]()
![]() 土 壤 学 报
土 壤 学 报
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]() high-effective resource utilization of crop straw residue, but also directly relates to the balance of soil carbon pool and its functionality. Despite a tremendous advance in the theoretical study on soil organic carbon priming following the external organic matter inputs, we are currently facing other big challenges. For instance, that how to combine this latest theoretical knowledge into the practical application of straw residue management aiming to promote soil carbon sequestration and reduce carbon emissions. To tackle these difficulties, the characteristics and potential mechanisms of the organic carbon priming due to straw return to soils should be clarified, and its specific drivers (i.e., edaphic, climatic and anthropogenic factors) should also be fully identified fully. This paper first systematically summarized the potential theoretical basis of organic carbon priming such as co-metabolism theory, N-mining theory, stoichiometric decomposition and microbial, and necromass reuse. Secondly, we combined existing research data and theoretical hypotheses to elaborate a series of implicit factors that govern the direction and magnitude of soil carbon priming, including the straw type and quantity, return method, water and fertilizer management, soil properties, and climatic agency, et al. Finally, taking into the consideration the high-effective utilization of straw residue, the sustainable build-up of soil carbon pool, and environmentally friendly farming practices, the potential research directions on straw-induced soil carbon priming we proposed, and the adaptative straw management methods designed to promote soil carbon sequestration were also outlined.
high-effective resource utilization of crop straw residue, but also directly relates to the balance of soil carbon pool and its functionality. Despite a tremendous advance in the theoretical study on soil organic carbon priming following the external organic matter inputs, we are currently facing other big challenges. For instance, that how to combine this latest theoretical knowledge into the practical application of straw residue management aiming to promote soil carbon sequestration and reduce carbon emissions. To tackle these difficulties, the characteristics and potential mechanisms of the organic carbon priming due to straw return to soils should be clarified, and its specific drivers (i.e., edaphic, climatic and anthropogenic factors) should also be fully identified fully. This paper first systematically summarized the potential theoretical basis of organic carbon priming such as co-metabolism theory, N-mining theory, stoichiometric decomposition and microbial, and necromass reuse. Secondly, we combined existing research data and theoretical hypotheses to elaborate a series of implicit factors that govern the direction and magnitude of soil carbon priming, including the straw type and quantity, return method, water and fertilizer management, soil properties, and climatic agency, et al. Finally, taking into the consideration the high-effective utilization of straw residue, the sustainable build-up of soil carbon pool, and environmentally friendly farming practices, the potential research directions on straw-induced soil carbon priming we proposed, and the adaptative straw management methods designed to promote soil carbon sequestration were also outlined.
Key words: Straw return; Priming effect; Soil organic carbon
土壤有机质分解的激发现象最早发现于 1926 年 Löhnis 开展的绿肥分解试验。1953 年, Bingeman 等[1]正式将该现象命名为“priming action or effect ”,
即在添加外源有机物质的土壤中, 土壤有机质的损 失较未添加的土壤有机质损失更大的现象。1963 年, 我国学者朱祖祥在绿肥肥效机制探讨中首次将激发 效应的概念引入我国, 并定义其为加入少量新鲜绿 肥而引起土壤原来所含难分解有机质和腐殖质突然 分解的现象[2]。目前, 使用更为广泛的是 Kuzyakov 等[3] 于 2000 年给出的定义,即由各种新鲜有机质添 加 引起土壤有 机质周转发 生强烈而短期改变的 现 象。激发效应按照有机质添加引起额外碳释放的来 源不同可将其分为表观激发和真实激发, 前者主要 源自微生物周转, 后者来自土壤有机质分解。按照 对土壤有机质分解速率改变的方向不同, 可分为正 激发与负激发[4]。
从 20 世纪 50 年代开始,国外研究人员利用同 位素标记技术对激发效应率先展开研究, 目前研究 内容已从最初论证这一现象的普遍性逐步发展到揭 示激发强度的变化过程及其驱动机制。科学界普遍 认为激发效应的产生主要是由于新鲜有机质输入引 起的微生物生物活性增强,进而导致土壤有机质分 解速率改变。就激发效应的发生机制,大量研究已 分别从共代谢、氮矿化、化学计量比和微生物残体 再利用等理论分别进行了探讨[5-8]。国内研究人员近
可溶性有机质等外源有机物质添加引起的激发效应 也进行了大量研究[5,9- 10]。同时, 利用室内培养[11]、 原位加热土壤和移植原状土柱模拟增温, 并结合整 合分析等多种方法[12] ,我国研究者从胞外酶活性、 微生物生物量等角度分析了激发效应的温度敏感性 机制,这进一步丰富了激发效应的理论研究。
作物秸秆是农田土壤有机碳库的重要外部补 充[13],其还田过程导致的激发效应普遍存在。尽管 大量研究已量化了秸秆引起的激发强度, 但在不同 研究中变异幅度较大, 主要归结于农田系统中激发 发生的多因子依赖性[14]。我国秸秆年产量居世界首 位[15],其资源化综合利用潜力巨大。秸秆还田是一 种简单易行增加土壤碳库的有效方式,近年来已得 到大面积的推广应用[16]。秸秆还田导致土壤有机碳 激发, 不仅关系到秸秆资源化利用效率, 而且涉及 农田土壤碳库构建。因此,系统阐明秸秆还田过程 对土壤有机质激发的驱动因素, 将对秸秆资源化高 效利用和农田土壤碳库科学管理具有重要意义。
据此, 本文首先对有机质分解激发效应发生的 可能机制进行了概述和系统总结。基于激发效应的 理论解析和数据集成, 进一步识别并论述了秸秆还 田过程中影响激发强度和方向的驱动因素。最后从 秸秆还田的高效性、农田碳库的可持续性和农业环 境的友好性出发, 对秸秆还田与有机碳耦合关系的 潜在研究方向进行了展望,并就优化秸秆还田改善
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]() 土壤碳库的管理措施提出了建议, 以期为我国秸秆
土壤碳库的管理措施提出了建议, 以期为我国秸秆
资源化高效利用提供理论支撑和技术参考。
1 秸秆还田下土壤有机质分解激发的 可能机制
秸秆还田土壤有机质激发效应是秸秆、微生物 以及土壤有机质共同参与的结果, 较多的研究已从 共代谢理论[17]、氮矿化理论[18]、化学计量比理论[19] 等方面来解释激发效应的强度和方向(图 1 )。
1.1 共代谢理论
共代谢理论可以理解为作物秸秆中易被分解的 简单碳源为土壤微生物提供初始的能量来源, 刺激 微生物体对特异性胞外酶的分泌, 尤其是参与降解 稳定性较高的秸秆组分的胞外酶[20] 。在此过程中, 与秸秆组成和结构相类似的稳定性较高的土壤有机 质同时被矿化而触发激发[21] ,Shahbaz 等[17]发现酶 活性和微生物数量与秸秆的分解相关,且微生物生 物量碳中土壤有机质源碳增加。该研究表明土壤碳 的矿化与秸秆添加引起的酶活性增强有关,且正激 发 的产生伴随 着微生物生 物量和胞外 酶的同步增 加。进一步分析主导土壤碳分解的微生物群落发现: 周转速度更快的“r ”型微生物首先降解秸秆易分解 碳源并合成胞外酶[22] ,增加的酶活性进而促进周转 速率较低的“k”型微生物对难分解的土壤有机质的 矿化分解。与之相反, 有研究发现:尽管真菌作为 典型的“k”型微生物, 在外源有机质输入后反而表 现出较“r ”型微生物更高的富集程度,且真菌体内 外源 13C 富集得更多[23]。可见, 基于共代谢理论的 “k”型和“r ”型微生物通过胞外酶分泌来主导激发 效应的机制仍需进一步研究。
1.2 氮矿化理论
氮矿化理论认为激发效应的发生是由秸秆碳输 入使微生物增长, 以及对氮素需求增加, 进而导致 微生物从有机质中矿化氮素引起的[24] 。换言之, 秸 秆输入导致土壤氮有效性在短期内降低, 且碳氮比 失衡, 这种环境下微生物需要矿化土壤有机质来获 取额外的氮素以满足生长和繁殖[25] 。由于氮素和碳 素的矿化是耦合的,氮的矿化伴随着碳的分解,土 壤 有 机 质 分 解 激 发 由 此 发 生 。 Moorhead 和 Sinsabaugh[26] 研究发现随养分有效性增加,激发强 度减弱。而输入较高碳氮比的秸秆更易导致土壤微
生物处于“氮饥饿”状态,因而更易增加激发强度[27-28]。 可见激发的发生与土壤中氮的有效性相关。尽管氮 矿化理论更多强调微生物氮素限制在引起激发效应 过程中的主导作用,然而微生物的生长和繁殖还离 不开对其他养分元素如磷,硫等的利用, 尤其是在 一些特定养分元素限制的区域。
1.3 化学计量比
化学计量比理论主张微生物活性受最稀缺的营 养元素的限制, 当养分供应与微生物的化学计量比相 匹配时, 微生物活性增加, 土壤有机碳矿化加快, 更 易导致激发的产生[17] 。该理论认为在养分计量比更 加平衡的状态下,微生物活性能被最大程度地激发 (尤其是“r ”型微生物)。因此, 当整个体系中养分 化学计量比符合微生物增长所需的最佳养分化学计 量比时, 微生物活性和胞外酶的分泌增加, 微生物对 土壤有机质分解加速, 从而产生激发。例如, 对水稻 秸秆分解及激发效应的研究表明: 氮磷共施相对更加 满足微生物化学计量需求, 因而促进土壤有机质的分 解[8] 。养分有效性较高的情况下,微生物群体较大, 更多受到呼吸能量的限制, 因此激发的碳可能更多来 自微生物的呼吸作用。而氮矿化理论支持激发的碳来 自于在养分相对亏缺时,部分活跃的微生物为了 “攫 取”土壤有机质中的养分而产生的“副产物”这一观 点, 同时这部分养分更多的是用于微生物构建自身生
命体。虽然该理论与氮矿化理论观点存在差异, 但有 研究认为这两种理论可能分别解释土壤有机碳矿化 的不同阶段激发产生的原因[19]。
1.4 微生物残体再利用
大量外源碳输入, 微生物快速摄取能量用于构 建自身生物量(合成代谢)的同时,自身代谢产物 积累,微生物对这些代谢产物的再利用(分解代谢) 也随之加快。大量低质量秸秆输入通常驱动特定微 生物种群优势扩大[29] ,这些微生物分解相对稳定的 碳库同时, 通过“埋藏效应”在稳定的碳库中引入 了更多的残体物质,使其成为土壤有机质的一部分。 微生物体 C ︰ N 比值相对较低, 且各组分没有与矿 物质紧密结合,因此当过量的秸秆碳输入,导致土 壤 C ︰ N 极端失衡, 土壤有机质不能满足微生物对 氮的需求时,这时已累积的微生物残体和代谢产物 易被其他类群微生物利用,以克服化学计量比的不 平衡, 耦合的碳组分随之被分解, 造成土壤本底有 机碳的额外激发。 Shahbaz 等[17]从秸秆质量和数量
![]()
探究土壤碳激发发现: 土壤呼吸增加伴随着微生物 生物量的减少和特异性酶活性的增强,但未发现土 壤本底有机碳被整合到微生物体中,表明激发的碳
源可能来自微生物残体的再利用。由此可见, 微生 物残体既是形成稳定土壤有机质的重要基底, 又是 激发效应的底物。

注: a )共代谢理论; b)氮矿化理论; c )化学计量比理论与微生物残体再利用理论。 Note: a ) co-metabolism; b) N-mining;
c ) stoichiometric decomposition & microbial necromass reuse .
图 1 激发效应发生的理论过程模式图
Fig. 1 Theoretical process pattern diagram of priming effect
2 秸秆还田下土壤有机质分解激发的 影响因素
基于上述对秸秆添加引起土壤有机质分解激发 潜在机理的认识, 进一步甄别秸秆还田过程中导致 激发发生的生物和非生物因素,能为提升秸秆还田 的土壤固碳潜力提供科学指导和技术借鉴。
2.1 秸秆类型
不同作物来源的秸秆因其质量差异(如氮浓度、 木质素和可溶性组分含量、 C ︰ N 等),往往导致土 壤有机质激发强度不同。 Schmatz 等[30]研究发现: 野豌豆秸秆本身较高的可溶性碳组分能够为微生物 提供较多的活性碳源, 从而促进激发。可溶性组分 含量较高的秸秆, 其自身分解速率相对较快, 能够 更快释放其他养分并刺激微生物生长, 进而引起更强
秆分解后释放的 N 、S 含量较高[19] ,土壤有机质激 发更强。此外,秸秆 C ︰ N 通常与土壤碳激发存在 一定的关系: Yanni 等[32] 和 Maokui 等[33]发现土壤碳 矿化速率与秸秆 C ︰ N 呈负相关。因秸秆 C ︰ N 远 大于微生物体 C ︰ N,富含碳的秸秆输入会造成土壤 微生物的养分限制。而较低 C ︰ N 的秸秆输入, 通 过提供更易利用的养分,能够更大程度地刺激微生 物活性,在此过程中, 土壤有机质中不易被利用的 组分也因微生物活性增加而被分解和利用[32] ,最终 导致土壤碳激发增强。 然而, 另有研究表明: 相比
大豆秸秆,C ︰ N 较高的玉米秸秆还田后会迫使微生 物(尤其是 k 策略型) 从矿物结合态有机质中矿化 更多的氮素来缓解化学计量比的失衡,最终导致更
强的有机质激发[34] 。本研究整合分析发现:尽管小
麦和玉米秸秆添加的土壤有机质激发强度无显著差
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]() 的激发[31] 。有研究表明: 与小麦秸秆相比,油菜秸 异,但玉米和小麦秸秆还田造成的激发强度显著高
的激发[31] 。有研究表明: 与小麦秸秆相比,油菜秸 异,但玉米和小麦秸秆还田造成的激发强度显著高
于豆科和水稻来源的秸秆(图 2 ),这一趋势符合秸 秆 C ︰ N 与激发强度呈正相关的结论。尽管大量的 研究已量化了秸秆 C ︰ N 与激发强度的关系,但仍 有研究发现秸秆的化学组成(如酚类、非结构性碳 水化合物等) 相比秸秆 C ︰ N 在影响有机质激发中 更具主导作用。如 Liang 等[35] 的研究发现: 土壤微 生物群落结构并不能与秸秆 C ︰ N 建立一致的关系, 秸秆中木质素等其他化学组成可能会导致微生物群 落结构发生改变, 不同微生物菌群对秸秆和土壤有机 质的分解能力不同[36] ,因此秸秆的化学组成会影响 土壤有机质的激发强度[37] 。可见,影响有机质激发 强度的秸秆属性不仅涉及其活性底物的多少, 同时还 受限其化学组成的复杂程度, 未来研究需要关注土壤 微生物对不同秸秆及其不同组成物质的可利用能力。

图 2 不同类型作物秸秆的土壤有机质分解激发强度比较
Fig. 2 Comparisons of the priming intensity response to different
types of crop straw
2.2 耕作方式
作物秸秆还田引起的土壤有机质激发受耕作方 式(如传统翻耕、少、免耕等)的显著影响。 Bimüller 等[38] 和 Guppy 等[39]对比了少免耕与翻耕条件下的 秸秆还田, 发现后者导致了更为明显的土壤有机质 激发。一般而言, 频繁耕作对土壤团聚体造成物理 破坏, 使其结构稳定性变差。在秸秆分解过程中释 放的小分子有机酸使矿物保护的土壤有机质溶解, 被团聚体保护的有机碳更易暴露且被微生物利用, 有机碳的生物有效性提高[40] 。Sarker 等[31] 的研究进
一步表明这种团聚体有机碳可利用性的提高显著增 加其矿化, 并伴随着激发效应的增强。 此外, 耕作
强度还会影响土壤有机碳库组分, 尤其是不稳定碳 组分, 进而导致这部分有机质更易被激发。与免耕 相比, 翻耕秸秆还田使土壤活性碳库有效性提高, 促进细菌生长和活性增强,加速了微生物对有机碳
的分解激发[41] 。最近的研究发现:相同条件下翻耕
还田的土壤酶转化效率较低, 需要微生物释放更多 的胞外酶到土壤中,这种酶的增加使碳底物和胞外 酶结合的可能性提高,进而导致土壤碳激发增强[42]。 相应地,免耕处理下土壤容重增加,土壤透气性降 低,最终影响好氧微生物群落的增殖,减缓土壤有 机质矿化及其激发效应[43] 。Sauvadet 等[42]还发现: 相比翻耕, 免耕条件下的土壤微生物具有更高的碳 利用效率, 一定程度上微生物降低了对土壤有机质 的矿化分解并表现出较弱的激发效应。此外, 耕作 方式还会影响土壤本底有机碳含量,土壤有机碳含 量与激发强度也存在一定关系, 如 Sun 等[44] 的研究
结果显示土壤碳激发强度随土壤有机碳含量的增加
而下降 。本文整合分析结果表明: 碳激发与土壤有
机碳含量存在一定关系, 当土壤有机碳含量在 10~
20 g ·kg– 1 区间内激发强度最高, 高于或低于该阈值
时激发变弱 (图 3 )。以上这种关系可能源于秸秆
添加导致 处于土壤有 机碳含量较低环境中的微生 物更加快速增长, 微生物对原土壤有机碳分解程度 更高[20]。此外 ,土壤碳激发的难易程度与本底碳的
稳定性也有关系: 碳含量越高的土壤中碳的吸附电 位更多,本底碳的稳定性提升,土壤碳库抗分解能
力增强,因此土壤碳激发更弱 [45] 。然而, Dimassi
等[46] 的研究表明: 虽然耕作强度能够改变土壤有机 碳以及微生物生物量的含量, 但最终的激发强度主 要受限于土壤活性有机碳与养分含量的比值关系, 因此与耕作方式相比, 土壤中养分的有效性更能解 释有机质激发的强度。
2.3 秸秆还田量
秸秆还田量关系到微生物可利用碳底物的有效 性,最终影响对土壤有机质的激发。尽管大量的研 究表明激发强度与外源碳添加量呈正相关[47-48] ,但 Blagodatskaya 和 Kuzyakov[20] 的整合研究发现外源 碳输入量与激发强度存在非线性关系,即当秸秆碳 输入量超过土壤微生物生物量碳的 50%时, 激发强 度开始降低, 当超过微生物生物量碳的 2 倍~5 倍
![]()
![]()
![]()
![]() 土 壤 学 报
土 壤 学 报
时,激发强度趋于零或变为负值。另有研究也证实 了过量秸秆碳输入后激发效应的饱和现象[49] ,甚至
激发强度下降[50-51] 。本研究的综合分析同样表明:
秸秆碳输入量大于相当于 30%土壤有机碳含量时,
土壤有机质激发有减弱的趋势 (图 4 )。
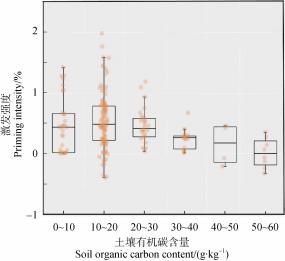
图 3 不同土壤有机碳含量等级下激发强度
Fig. 3 Priming intensity under different soil organic carbon content levels
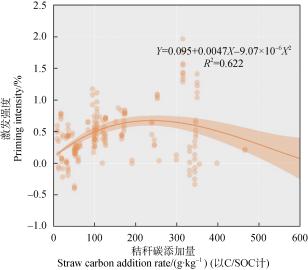
图 4 激发强度与秸秆碳添加量的关系
Fig. 4 Relationship between the priming intensity and the addition
rate of straw carbon
在相对低量的秸秆输入情况下,激发强度随秸 秆添加量的增加而增大,这主要与微生物群落结构 的变化和它对其他营养物质的需求有关。从养分需 求角度解释: 相对较高含量的秸秆输入, 使微生物获
得大量可利用组分, 激活并使微生物生物量增加[50],
此时微生物开始受到氮限制, 导致微生物开始矿化
土壤有机质中的氮而伴随碳激发[18] ;Blagodatskaya
和 Kuzyakov[20]从微观角度分析发现, 具有分解秸秆 和土壤碳功能微生物数量的变化, 可能会导致整个 土壤微生物群落结构发生变化,并认为与少量外源 基质添加相比,更多的基质添加首先会使土壤中部 分“获益”于秸秆养分的微生物活化,之后, 处于 休眠状态的特定微生物被激活,并且这些微生物可 以快速生长并成为优势物种, 在降解外源基质的过 程中促进了对土壤有机质的共代谢,因此有机碳的 激发增强。 Landi 等[52]通过凝胶电泳和磷脂脂肪酸 分析进一步验证了真实激发的增强伴随着微生物群 落结构的改变。相对应的较少量的秸秆添加, 没有 导致微生物量的大小和群落组成发生变化,且微生 物未释放更多的胞外酶去分解原土壤有机质, 故激 发强度较低。其次,过量秸秆输入可能导致土壤有 机质激发强度下降,主要有以下两个支持性观点:
大量研究表明, 这种秸秆大量输入后,微生物从利
用养分有效性较低的土壤有机质转向利用养分有效
激发“饱和”的现象与土壤有机质的可接触性有关, 尽管微生物可以产生大量的酶,但仍有部分有机质 被保护而构成稳定的碳库,许多有机化合物仍不能 被触及,因此激发强度被土壤碳库的稳定性限制[54], Falloon 和 Smith[55]研究显示稳定而不易被分解的有 机碳约占土壤总有机碳的 15%~59%,这部分有机 碳的存在导致激发强度逐渐平稳。
2.4 肥料管理
施肥措施直接决定土壤养分的有效性,进而改 变土壤微生物对底物的利用情况, 最终影响土壤有 机质的激发强度。 Wang 等[56]通过室内培养发现: 无机氮添加提高了土壤氮素有效性,降低了微生物 对土壤有机质中氮的获取,最终显著降低了土壤碳 的激发强度,该研究也一定程度上解释了在土壤养 分含量相对较低的农田进行秸秆还田后土壤碳激发
增强的现象。 在氮素有效性较低的情况下,大量秸 秆输入导致土壤底物中碳浓度增加,加剧了微生物 的养分限制,迫使微生物体从土壤有机质中矿化出 更多的 N 、P 、S 等养分元素来克服化学计量比不平 衡性以保证自身的生长和繁殖,有机质中养分的释 放和有机碳的矿化是耦合的, 额外的养分获取伴随
![]()
着有机碳分解的激发[57] 。氮素有效性的改善通常能 够显著降低土壤有机质的激发强度。一方面, 解除 微生物的氮限制, 另一方面, 氮素添加后导致土壤 溶液中有毒物质渗透, 微生物量下降,微生物对土 壤有机质的矿化能力减弱[46] ,最终导致激发弱于低 养分处理的土壤碳激发。然而, 另有研究发现土壤养 分有效性较高触发的有机质激发强度反而更大[58] 。 依据 Kuzyakov 和 Bol[59] 的研究: 激发强度与土壤 有机碳含量呈正相关, 微生物活性随碳含量增加而 升高。当在较高土壤养分条件下进行秸秆还田时, 土壤有机碳积累和微生物活性的增加[60] ,很可能导 致更大的有机质激发。由此可见, 养分有效性与碳 激发关系的中间因子可能是土壤有机碳含量与微生 物活性。从微生物群落组成角度来看,高养分环境 意味着化学计量比( C ︰ N ︰ P)较低,可能更利于 专性降解稳定性高的土壤有机质的微生物占据主导 地位, 这种微生物在高碳、高氮的环境下持续专性 分解土壤有机质, 促进了土壤碳的激发[19]。进一步, Fang 等[21] 和 Chen 等[19]从碳矿化的时间尺度上研究 发现土壤高养分有效性触发激发通常发生在秸秆分 解的后期[19,21]。此外, 分解有机物的微生物活性的增 加除了与土壤氮有效性有关外, 还与磷元素有关[61], 如土壤中添加无机磷缓解微生物的磷限制 [62] 并与 吸附的有机化合物发生交换, 使有机化合物分解增 强[63] ,最终增强了土壤碳激发。基于以上研究结果 和作物生产需要,建议秸秆还田前期合理配施化肥, 改善土壤养分状况,使土壤处于养分均衡状态,并 减少土壤有机碳矿化[64] 。将来的研究需进一步探究 外源养分资源输入与微生物需求之间的平衡问题, 并结合土壤碳矿化,更加全面、系统地评估秸秆还 田与肥料配施对土壤肥力提升的效应。
2.5 温度
温度是一切酶促反应的重要驱动[65] ,酶活性的 改 变是变温过 程中土壤有 机碳分解的 主要驱动 因 子。偏离酶活性最适温度的增温或者降温均会影响 土壤酶对新鲜有机质的分解以及对本底有机碳的矿 化分解,进而影响激发效应的强度和方向[66] 。生产 实践中,地理位置、栽培方式、水分条件等因素的 不同直接作用于土壤温度状况[67-68] ,这很可能最终 影响秸秆还田引起的土壤有机质激发效应。通常认 为: 相对于低温条件, 温度升高可降低反应活化能,
提高酶的反应速率,土壤的呼吸速率上升[69-70] 。温 度敏感性 Q10 (温度每增加 10℃土壤呼吸所增加的 倍数) 经常被用来衡量变温对土壤碳矿化的影响。 研究发现: 与低温相比,持续升温导致激发效应的 温度敏感性逐渐下降[71] 。Liu 等[72] 的研究显示:胞 外酶活性与土壤有机碳源 CO2 排放速率呈正线性相 关,但随着时间延长, 酶活性随温度升高而降低, 导致激发效应温度敏感性(Q10 )与温度呈负相关[73]。 从反应底物的有效性来看,温度逐渐升高使土壤变 得更加干燥, 外源可溶性基质溶解、扩散速率降低, 微生物可利用的碳源减少,激发效应的温度敏感性 下降[74] 。在秸秆分解后期,有机质的分解对增温的 响应逐渐减弱,可能归因于微生物对升温的逐渐适 应或发生了酶降解[72,75] 。此外,有研究发现土壤碳 矿化的温度敏感性还与土壤碳库的质量有关: 室内 培养长期原位加热的森林土壤发现,土壤木质素等 抗分解的物质含量较高,导致土壤中的微生物可利 用养分的有效性下降, 微生物种群数量受到限制, 土壤碳的分解速率降低,因此仅提升微生物活动的 温度不会刺激土壤碳的分解[76] ,最终这种土壤碳分 解速率的减小导致累积土壤碳激发减弱。尽管激发 效应的发 生是由微生 物群落的特定组成部分 主导 的[77],但温度对土壤微生物群落结构的长期影响以 及由此驱动的土壤碳的激发还未被完全了解, 它们 之间的关系尚需进一步研究。
2.6 水分
水分状况决定了可溶性底物在土壤中的扩散程 度,从而影响微生物体对底物的利用程度,最终改 变土壤有机质的激发强度。大量研究借助干湿交替 的室内模拟已探讨了水分有效性对土壤有机质激发 效应的影响,这为不同气候条件下秸秆还田引起的 激发强度的研究提供了重要借鉴。研究表明: 与持 续湿润的土壤相比,秸秆添加后干湿循环处理使土 壤碳激发高出 1.71~3.58( mg ·kg– 1 · d– 1,以 C 计) [78]。 土壤水分过高,微生物遭受缺氧可能导致其活性下 降。与持续干燥的土壤相比, 干湿循环使土壤碳排 放增加约 72%[79]。土壤水分有效性很大程度上决定 可溶性养分的扩散程度,影响养分的可利用性,土 壤水分过低时土壤表面水膜厚度减小,基质向微生 物的扩散速度降低[80] ,秸秆添加后微生物可利用养 分有效性低,导致土壤碳激发的反应速率下降。一
![]()
方面, 干湿循环会导致自由水快速摄入, 空气被截 留在孔隙中,再经历冻融使土壤团聚体和微生物经 历物理损伤,这种膨胀破坏可能导致大团聚体周转 加速以及其中有机物损失[81] ,这种团聚体结构被破 坏后有机物质被释放的这部分土壤碳更易被激发。 经历干燥时部分微生物死亡, 土壤复湿后氨基酸、 甘油等细胞溶质暴露增加并被微生物再利用, 微生 物可利用养分的增加促进了它对土壤本底碳的分解 能力, 这个过程中微生物残体和秸秆分解的副产物 均参与土壤碳激发增强的过程。另一方面,干湿循 环还会影响土壤的通气性,创造交替的厌氧和好氧 环境, 使微生物种类更加丰富,尤其是细菌群落的 变化可以解释部分地区土壤经历干湿循环后土壤碳 矿化速率的下降[82] ,土壤碳激发由微生物驱动,若 微生物对碳的矿化速率下降, 也就意味着对土壤本 底碳的激发有所缓解。部分研究者认为大多数团聚 体在经历了干湿循环后会逐渐适应湿润环境, 有机 碳在大团聚体中仍处于被保护状态[83] ,对微生物的 分解仍具有物理防护作用,这种不可接触性保护有 机碳不易被激发。干燥或湿润持续时间和循环次数 与微生物能否适应环境压力、保持活性或面临死亡 存在关联,所以可能造成激发强度存在差异。 Zhang 等[79]通过整合分析发现在干湿循环次数较多的土壤 中,团聚体的物理保护作用在较稳定土壤碳的分解 过程起着决定性作用, 而土壤本底碳库能否经受多 次循环,保持团聚体结构不被破坏这些问题均需要 更进一步探究。
3 结论与展望
土壤有机质是农田肥力的基础与核心,对作物 产量和农业环境可持续性具有重要意义。作物秸秆 作为农田土壤有机碳库的重要外部来源, 对改善土 壤有机质状况具有显著影响。秸秆还田导致的土壤 有机质分解的激发效应是普遍存在的,但其发生机 制是复杂的。总体而言,激发效应的发生和动态变 化主要是由土壤微生物体对不同来源的底物进行差 异化利用而造成的。就土壤有机质本身而言, 其不 同的稳定程度是决定激发强度的重要因素。激发效 应发生的方向和强度是多变的,但秸秆还田对农田 土壤有机碳的影响缓慢且深远,由于作用时间的滞
后性, 目前还未将秸秆还田下土壤有机质分解激发 与秸秆腐解过程中能量和养分释放建立直接联系。 因此探究长期土壤有机碳输入与输出的动态平衡将 对优化秸秆还田技术具有宏观指导意义。
我国农业气候资源多样,且各地水热条件和农 业管理模式差别较大, 导致秸秆还田对土壤有机质 激发的强度不同, 探究不同地区环境因子和人为因 素对激发引起的碳损失的相对贡献,对农田土壤碳 库管理具有重要的实践意义。但考虑到秸秆还田土 壤有机质分解激发的普遍性及其对土壤碳库影响的 显著性,实施秸秆还田技术需要关注土壤有机质分 解激发强度和方向的影响因素。比如长期单一作物 秸秆还田不利于构建多样的微生物环境, 易造成同 一营养型微生物富集,对土壤有机碳造成长期损耗, 需要结合不同 地区光热资 源发展多种作物配套 还 田;减少长期深翻对土壤有机质物理保护屏障造成 的破坏,需要塑造良好的土壤微环境,改进秸秆还 田技术以适应不同作物及耕作制度,在土壤质地较 好且适宜发展保护性农业的地区推广秸秆覆盖和翻 耕还田相结合的还田策略;结合区域土壤养分水平 和微生物组成与结构特征,不同地区需要考虑还田 量及秸秆类型来设计还田策略;土壤微生物不仅是 土壤有机物质转化的执行者, 又是植物营养元素的 活性库。要充分发挥微生物的调节功能, 调节施肥 配比及施用时间, 构建秸秆腐解微生物和作物间的 养分平衡利用关系;减小环境因子对土壤有机碳损 失的影响, 需要根据气候条件调节秸秆还田时间, 实现秸秆资源的更高效利用。
参考文献( References )
[ 1 ] Bingeman C W,Varner J E,Martin W P. The effect of the
addition of organic materials on the decomposition of an organic soil[J] . Soil Science Society of America Journal, 1953 , 17 ( 1 ): 34—38 .
[ 2 ] Zhu Z X . Discussing the fertilizer effect mechanism of
green manure and its application problems[J]. Journal of Zhejiang Agricultural Sciences,1963,4 ( 3 ):104— 109 . [朱祖祥 . 从绿肥的起爆效应探讨它的肥效机制及其在 施用上的若干问题[J]. 浙江农业科学, 1963 ,4 ( 3 ):
104— 109 .
[ 3 ] Kuzyakov Y , Friedel J K , Stahr K. Review of
mechanisms and quantification of priming effects[J] . Soil Biology and Biochemistry , 2000 , 32 ( 11/12 ):
1485— 1498 .
![]()
![]() 6 期 张叶叶等:秸秆还田下土壤有机质激发效应研究进展 1389
6 期 张叶叶等:秸秆还田下土壤有机质激发效应研究进展 1389
![]()
![]() Blagodatsky S ,Blagodatskaya E ,Yuyukina T ,et al . Model of apparent and real priming effects: Linking microbial activity with soil organic matter decomposition[J] . Soil Biology and Biochemistry,2010, 42 ( 8 ): 1275— 1283 .
Blagodatsky S ,Blagodatskaya E ,Yuyukina T ,et al . Model of apparent and real priming effects: Linking microbial activity with soil organic matter decomposition[J] . Soil Biology and Biochemistry,2010, 42 ( 8 ): 1275— 1283 .
![]()
![]() Cui J ,Zhu Z K ,Xu X L ,et al . Carbon and nitrogen recycling from microbial necromass to cope with C ︰ N stoichiometric imbalance by priming[J]. Soil Biology and Biochemistry ,2020 , 142: 107720.
Cui J ,Zhu Z K ,Xu X L ,et al . Carbon and nitrogen recycling from microbial necromass to cope with C ︰ N stoichiometric imbalance by priming[J]. Soil Biology and Biochemistry ,2020 , 142: 107720.
![]()
![]() Jenkins S N , Rushton S P , Lanyon C V , et al . Taxon-specific responses of soil bacteria to the addition of low level C inputs[J] . Soil Biology and Biochemistry, 2010 ,42 ( 9 ): 1624— 1631 .
Jenkins S N , Rushton S P , Lanyon C V , et al . Taxon-specific responses of soil bacteria to the addition of low level C inputs[J] . Soil Biology and Biochemistry, 2010 ,42 ( 9 ): 1624— 1631 .
![]()
![]()
![]()
![]() Li L J ,Zhu-Barker X ,Ye R Z ,et al . Soil microbial biomass size and soil carbon influence the priming effect from carbon inputs depending on nitrogen availability[J]. Soil Biology and Biochemistry ,2018 , 119 :41—49 . Zhu Z K,Ge T D,Luo Y,et al . Microbial stoichiometric flexibility regulates rice straw mineralization and its priming effect in paddy soil[J]. Soil Biology and Biochemistry ,2018 , 121: 67—76 .
Li L J ,Zhu-Barker X ,Ye R Z ,et al . Soil microbial biomass size and soil carbon influence the priming effect from carbon inputs depending on nitrogen availability[J]. Soil Biology and Biochemistry ,2018 , 119 :41—49 . Zhu Z K,Ge T D,Luo Y,et al . Microbial stoichiometric flexibility regulates rice straw mineralization and its priming effect in paddy soil[J]. Soil Biology and Biochemistry ,2018 , 121: 67—76 .
![]()
![]() Li Y ,Zhou C F ,Qiu Y X ,et al . Effects of biochar and litter on carbon and nitrogen mineralization and soil microbial community structure in a China fir plantation[J] . Journal of Forestry Research,2019,30( 5 ):
Li Y ,Zhou C F ,Qiu Y X ,et al . Effects of biochar and litter on carbon and nitrogen mineralization and soil microbial community structure in a China fir plantation[J] . Journal of Forestry Research,2019,30( 5 ):
1913— 1923 .
![]()
![]() Yuan S F ,Wang S L ,Zhang W D . Effect of external organic carbon and temperature on SOC decomposition[J] . Chinese Journal of Soil Science,2015, 46 ( 4 ) :916—922 . [袁淑芬, 汪思龙, 张伟东 . 外源有 机碳和温度对土壤有机碳分解的影响[J]. 土壤通报, 2015 ,46 ( 4 ): 916—922 .]
Yuan S F ,Wang S L ,Zhang W D . Effect of external organic carbon and temperature on SOC decomposition[J] . Chinese Journal of Soil Science,2015, 46 ( 4 ) :916—922 . [袁淑芬, 汪思龙, 张伟东 . 外源有 机碳和温度对土壤有机碳分解的影响[J]. 土壤通报, 2015 ,46 ( 4 ): 916—922 .]
![]()
![]() Yuan S F ,Wang S L ,Zhang W D . Effect of external organic carbon and temperature on soc decomposition . Chinese Journal of Soil Science ,2015.
Yuan S F ,Wang S L ,Zhang W D . Effect of external organic carbon and temperature on soc decomposition . Chinese Journal of Soil Science ,2015.
![]()
![]() Zhang W D ,Wang X F ,Wang S L . Addition of external organic carbon and native soil organic carbon decomposition:A meta-analysis[J] . PLoS One ,2013 , 8 ( 2 ): e54779 .
Zhang W D ,Wang X F ,Wang S L . Addition of external organic carbon and native soil organic carbon decomposition:A meta-analysis[J] . PLoS One ,2013 , 8 ( 2 ): e54779 .
![]()
![]()
![]()
![]()
![]() Jin Z Q,Shah T,Zhang L,et al . Effect of straw returning on soil organic carbon in rice-wheat rotation system :A review[J] . Food and Energy Security,2020,9 ( 2 ):e200 . Ding F , van Zwieten L , Zhang W D , et al . A meta-analysis and critical evaluation of influencing factors on soil carbon priming following biochar amendment[J]. Journal of Soils and Sediments,2018 ,18 ( 4 ): 1507— 1517 .
Jin Z Q,Shah T,Zhang L,et al . Effect of straw returning on soil organic carbon in rice-wheat rotation system :A review[J] . Food and Energy Security,2020,9 ( 2 ):e200 . Ding F , van Zwieten L , Zhang W D , et al . A meta-analysis and critical evaluation of influencing factors on soil carbon priming following biochar amendment[J]. Journal of Soils and Sediments,2018 ,18 ( 4 ): 1507— 1517 .
![]()
![]() Jiao L N ,Li H Z ,Yin C C . Research status of straw returning in improving soil fertility[J] . Anhui Agricultural Science Bulletin ,2014 ,20 ( 10 ) :54— 56 .
Jiao L N ,Li H Z ,Yin C C . Research status of straw returning in improving soil fertility[J] . Anhui Agricultural Science Bulletin ,2014 ,20 ( 10 ) :54— 56 .
[矫丽娜, 李志洪,殷程程 . 秸秆还田培肥土壤的研究
![]()
![]()
![]()
![]() 现状[J]. 安徽农学通报, 2014 ,20 ( 10 ): 54— 56 .]
现状[J]. 安徽农学通报, 2014 ,20 ( 10 ): 54— 56 .]
![]()
Dou L Y. Effects of straw return to field on stability , organic carbon content and distribution of different types of soil aggregates[D] . Shenyang: Shenyang Agricultural University ,2018. [窦莉洋 . 秸秆还田对不同类型土壤 团聚体稳定性、有机碳含量及其分布的影响[D] . 沈阳: 沈阳农业大学, 2018 .]
Shahbaz M,Kuzyakov Y,Sanaullah M,et al . Microbial decomposition of soil organic matter is mediated by quality and quantity of crop residues :Mechanisms and thresholds[J] . Biology and Fertility of Soils ,2017 ,53 ( 3 ) :287—301 .
Fontaine S ,Henault C ,Aamor A ,et al . Fungi mediate long term sequestration of carbon and nitrogen in soil through their priming effect[J]. Soil Biology and Biochemistry ,2011 ,43 ( 1 ): 86—96 .
Chen R R,Senbayram M,Blagodatsky S,et al. Soil C and N availability determine the priming effect :Microbial N mining and stoichiometric decomposition theories[J] . Global Change Biology ,2014 ,20 ( 7 ) :2356—2367 . Blagodatskaya Е ,Kuzyakov Y. Mechanisms of real and apparent priming effects and their dependence on soil microbial biomass and community structure : Critical review[J] . Biology and Fertility of Soils ,2008 ,45 ( 2 ):
115— 131 .
Fang Y Y ,Nazaries L , Singh B K ,et al . Microbial mechanisms of carbon priming effects revealed during the interaction of crop residue and nutrient inputs in contrasting soils[J] . Global Change Biology ,2018 ,24 ( 7 ) :2775—2790 .
Schimel J P , Bennett J . Nitrogen mineralization : Challenges of a changing paradigm[J]. Ecology ,2004, 85 ( 3 ): 591—602 .
Fabian J ,Zlatanovic S ,Mutz M ,et al . Fungal-bacterial dynamics and their contribution to terrigenous carbon turnover in relation to organic matter quality[J] . The ISME Journal ,2017 , 11 ( 2 ) :415—425 .
Craine J M ,Morrow C ,Fierer N. Microbial nitrogen limitation increases decomposition. Ecology,2007,88( 8 ):
2105—2113.
Shahbaz M ,Kumar A ,Kuzyakov Y ,et al. Priming effects induced by glucose and decaying plant residues on SOM decomposition :A three-source 13C/14C partitioning study[J]. Soil Biology and Biochemistry ,2018 , 121: 138— 146. Moorhead D L , Sinsabaugh R L . A theoretical model of litter decay and microbial interaction[J] . Ecological Monographs ,2006 ,76 ( 2 ): 151— 174 .
Kuzyakov Y. Priming effects:Interactions between living and dead organic matter[J] . Soil Biology and Biochemistry ,2010 ,42 ( 9 ): 1363— 1371 .
Wild B,Li J,Pihlblad J,et al . Decoupling of priming and microbial N mining during a short-term soil incubation[J] .
![]()
![]() Soil Biology and Biochemistry ,2019,
Soil Biology and Biochemistry ,2019,![]() 129
129![]() :71—79 .
:71—79 .
1390
土 壤 学 报
58 卷
[ 29 ] Kaiser C ,Franklin O ,Dieckmann U ,et al . Microbial mineralisation and nutrient supply in contrasting soils
community dynamics alleviate stoichiometric constraints under long-term farming systems[J] . Soil and Tillage
during litter decay[J]. Ecology Letters ,2014 , 17 ( 6 ): Research ,2019 , 193: 71— 84 .
680—690 . [ 42 ] Sauvadet M ,Lashermes G ,Alavoine G , et al. High
[ 30 ] Schmatz R,Recous S,Aita C,et al . Crop residue quality carbon use efficiency and low priming effect promote soil
and soil type influence the priming effect but not the fate C stabilization under reduced tillage[J] . Soil Biology and
of crop residue C[J]. Plant and Soil ,2017 ,414 ( 1/2 ): Biochemistry ,2018 , 123: 64—73 .
229—245 . [ 43 ] Kan Z R,Virk A L,Wu G,et al. Priming effect intensity
[ 31 ] Sarker J R , Singh B P ,Cowie A L ,et al . Carbon and of soil organic carbon mineralization under no-till and
nutrient mineralisation dynamics in aggregate-size residue retention[J] . Applied Soil Ecology ,2020 , 147:
classes from different tillage systems after input of canola 103445.
and wheat residues[J] . Soil Biology and Biochemistry , [ 44 ] Sun Z L,Liu S G,Zhang T,et al . Priming of soil organic
2018 , 116 :22—38 . carbon decomposition induced by exogenous organic
[ 32 ] Lyu M K,Nie Y Y,Giardina C P,et al. Litter quality and carbon input :A meta-analysis[J] . Plant and Soil ,2019,
site characteristics interact to affect the response of 443 ( 1/2 ) :463—471 .
priming effect to temperature in subtropical forests[J] . [ 45 ] Zimmerman A R,Gao B,Ahn M Y. Positive and negative
Functional Ecology ,2019 ,33 ( 11 ) :2226—2238 . carbon mineralization priming effects among a variety of
[ 33 ] Yanni S F,Whalen J K,Simpson M J,et al . Plant lignin biochar-amended soils[J]. Soil Biology and
and nitrogen contents control carbon dioxide production Biochemistry ,2011 ,43 ( 6 ): 1169— 1179 .
and nitrogen mineralization in soils incubated with Bt [ 46 ] Dimassi B,Mary B,Fontaine S,et al . Effect of nutrients
and non-Bt corn residues[J] . Soil Biology and availability and long-term tillage on priming effect and
Biochemistry ,2011 ,43 ( 1 ): 63—69 . soil C mineralization[J] . Soil Biology and Biochemistry,
[ 34 ] Mazzilli S R ,Kemanian A R ,Ernst O R ,et al . Priming 2014 ,78: 332—339 .
of soil organic carbon decomposition induced by corn [ 47 ] Fontaine S,Mariotti A,Abbadie L . The priming effect of
compared to soybean crops[J] . Soil Biology and organic matter :A question of microbial competition?[J].
Biochemistry ,2014 ,75 :273—281 . Soil Biology and Biochemistry,2003,35 ( 6 ):837— 843 .
[ 35 ] Liang X,Yuan J,Yang E,et al . Responses of soil organic [ 48 ] Tian J,Pausch J,Yu G R,et al . Aggregate size and their
carbon decomposition and microbial community to the disruption affect 14C-labeled glucose mineralization and
addition of plant residues with different C ︰ N ratio[J]. priming effect[J] . Applied Soil Ecology,2015,90:1— 10 .
European Journal of Soil Biology ,2017 , 82: 50— 55 . [ 49 ] Paterson E , Sim A . Soil-specific response functions of
[ 36 ] Meidute S , Demoling F , Bååth E . Antagonistic and organic matter mineralization to the availability of labile
synergistic effects of fungal and bacterial growth in soil carbon[J] . Global Change Biology , 2013 , 19 ( 5 ):
after adding different carbon and nitrogen sources[J]. Soil 1562— 1571 .
Biology and Biochemistry,2008,40 ( 9 ):2334—2343 . [ 50 ] Blagodatskaya E V ,Blagodatsky S A ,Anderson T H,
[ 37 ] Hamer U,Marschner B . Priming effects of sugars,amino et al . Priming effects in Chernozem induced by glucose
acids,organic acids and catechol on the mineralization of and N in relation to microbial growth strategies[J] .
lignin and peat[J]. Journal of Plant Nutrition and Soil Applied Soil Ecology ,2007 ,37 ( 1/2 ): 95— 105 .
Science ,2002 , 165 ( 3 ) :261—268 . [ 51 ] Qiao N , Schaefer D ,Blagodatskaya E ,et al . Labile
[ 38 ] Bimüller C ,Kreyling O ,Kölbl A ,et al . Carbon and carbon retention compensates for CO2 released by
nitrogen mineralization in hierarchically structured priming in forest soils[J] . Global Change Biology,2014,
aggregates of different size[J] . Soil and Tillage Research , 20 ( 6 ): 1943— 1954 .
2016 , 160 :23—33 . [ 52 ] Landi L ,Valori F ,Ascher J ,et al . Root exudate effects
[ 39 ] Guppy C N , Menzies N W , Moody P W , et al . on the bacterial communities ,CO2 evolution ,nitrogen
Competitive sorption reactions between phosphorus and transformations and ATP content of rhizosphere and bulk
organic matter in soil:A review[J] . Soil Research,2005 , soils[J] . Soil Biology and Biochemistry ,2006 ,38 ( 3 ):
43 ( 2 ): 189—202 . 509— 516 .
[ 40 ] Xue J F ,Pu C ,Zhao X ,et al . Changes in soil organic [ 53 ] Kuzyakov Y. Review : Factors affecting rhizosphere
carbon fractions in response to different tillage practices priming effects[J]. Journal of Plant Nutrition and Soil
under a wheat-maize double cropping system[J] . Land Science ,2002 , 165 ( 4 ): 382.
Degradation & Development,2018,29( 6 ):1555— 1564 . [ 54 ] Lützow M V ,Kögel-Knabner I ,Ekschmitt K ,et al.
[ 41 ] Sarker J R , Singh B P ,Fang Y Y ,et al . Tillage history Stabilization of organic matter in temperate soils :
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]() and crop residue
and crop residue![]() input enhanced native
input enhanced native ![]() carbon
carbon ![]() Mechanisms
Mechanisms ![]() and
and ![]() their
their![]() relevance under
relevance under ![]() different
different![]()
![]() soil
soil
![]() 6 期 张叶叶等:秸秆还田下土壤有机质激发效应研究进展 1391
6 期 张叶叶等:秸秆还田下土壤有机质激发效应研究进展 1391
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
conditions - A review[J] . European Journal of Soil [ 67 ] Lenka S,Trivedi P,Singh B,et al . Effect of crop residue
Science ,2006 ,57 ( 4 ) :426—445 . addition on soil organic carbon priming as influenced by
[ 55 ] Falloon P D , Smith P. Modelling refractory soil organic temperature and soil properties[J] . Geoderma,2019,347:
matter[J] . Biology and Fertility of Soils,2000,30 ( 5/6 ): 70—79 .
388—398 . [ 68 ] Yu Y Y ,Turner N C ,Gong Y H ,et al . Benefits and
[ 56 ] Wang H ,Hu G Q ,Xu W H ,et al . Effects of nitrogen limitations to straw- and plastic-film mulch on maize
addition on soil organic carbon mineralization after maize yield and water use efficiency :A meta-analysis across
stalk addition[J]. European Journal of Soil Biology , hydrothermal gradients[J] . European Journal of
2018 , 89: 33—38 . Agronomy ,2018 ,99: 138— 147 .
[ 57 ] Fontaine S ,Barot S . Size and functional diversity of [ 69 ] Blagodatskaya E ,Blagodatsky S ,Khomyakov N ,et al.
microbe populations control plant persistence and Temperature sensitivity and enzymatic mechanisms of
long-term soil carbon accumulation[J]. Ecology Letters , soil organic matter decomposition along an altitudinal
2005 , 8 ( 10 ): 1075— 1087 . gradient on Mount Kilimanjaro[J] . Scientific Reports,
[ 58 ] Xu Y D,Ding F,Gao X D,et al . Mineralization of plant 2016 ,6 :22240.
residues and native soil carbon as affected by soil fertility [ 70 ] Daniel R M ,Danson M J . A new understanding of how
and residue type[J]. Journal of Soils and Sediments , temperature affects the catalytic activity of enzymes[J] .
2019 , 19 ( 3 ): 1407— 1415 . Trends in Biochemical Sciences , 2010 , 35 ( 10 ):
[ 59 ] Kuzyakov Y,Bol R. Sources and mechanisms of priming 584— 591 .
effect induced in two grassland soils amended with slurry [ 71 ] Melillo J M,Steudler P A,Aber J D,et al . Soil warming
and sugar[J]. Soil Biology and Biochemistry ,2006 ,38 and carbon-cycle feedbacks to the climate system[J] .
( 4 ): 747—758 . Science ,2002 ,298 ( 5601 ) :2173—2176 .
[ 60 ] Jin X X,Gall A R,Saeed M F,et al . Plastic film mulching [ 72 ] Liu Q Y , Xu X L , Wang H M , et al. Dominant
and nitrogen fertilization enhance the conversion of extracellular enzymes in priming of SOM decomposition
newly-added maize straw to water-soluble organic depend on temperature[J] . Geoderma , 2019 , 343 :
carbon[J] . Soil and Tillage Research,2020,197:104527. 187— 195 .
[ 61 ] Ilstedt U , Singh S . Nitrogen and phosphorus [ 73 ] Peterson M E ,Daniel R M ,Danson M J ,et al . The
limitations of microbial respiration in a tropical dependence of enzyme activity on temperature :
phosphorus -fixing acrisol ( ultisol ) compared with Determination and validation of parameters[J] . The
organic compost[J] . Soil Biology and Biochemistry , Biochemical Journal ,2007 ,402 ( 2 ): 331—337 .
2005 , 37 ( 7 ): 1407— 1410 . [ 74 ] Wang C K ,Yang J Y ,Zhang Q Z . Soil respiration in six
[ 62 ] Li Y,Niu S L,Yu G R . Aggravated phosphorus limitation temperate forests in China[J] . Global Change Biology,
on biomass production under increasing nitrogen 2006 , 12 ( 11 ) :2103—2114 .
loading :A meta-analysis[J]. Global Change Biology , [ 75 ] Alvarez G , Shahzad T ,Andanson L ,et al . Catalytic
2016 ,22 ( 2 ): 934—943 . power of enzymes decreases with temperature : New
[ 63 ] Spohn M , Schleuss P M. Addition of inorganic insights for understanding soil C cycling and microbial
phosphorus to soil leads to desorption of organic ecology under warming[J] . Global Change Biology ,
compounds and thus to increased soil respiration[J] . Soil 2018 ,24 ( 9 ) :4238—4250 .
Biology and Biochemistry ,2019 , 130 :220—226 . [ 76 ] Giardina C P,Ryan M G. Evidence that decomposition
[ 64 ] Ma X ,Wei L ,Tang M L ,et al. Effects of varying rates of organic carbon in mineral soil do not vary
long-term fertilization on organic carbon mineralization with temperature[J] . Nature , 2000 , 404 ( 6780 ):
and priming effect of paddy soil[J] . Environmental 858— 861 .
Science ,2018 ,39 ( 12 ): 5680— 5686 . [马欣, 魏亮, [ 77 ] Romaní A M , Fischer H , Mille-Lindblom C , et al .
唐美玲, 等 . 长期不同施肥对稻田土壤有机碳矿化及激 Interactions of bacteria and fungi on decomposing litter:
发效应 的影 响 [J]. 环 境 科 学 , 2018 , 39 ( 12 ) : Differential extracellular enzyme activities[J] . Ecology,
5680— 5686 .] 2006 , 87 ( 10 ) :2559—2569 .
[ 65 ] Davidson E A ,Janssens I A . Temperature sensitivity of [ 78 ] Liu E K ,Wang J B ,Zhang Y Q ,et al. Priming effect of
soil carbon decomposition and feedbacks to climate ( 13 ) C-labelled wheat straw in no-tillage soil under
change[J]. Nature ,2006 ,440 ( 7081 ): 165— 173 . drying and wetting cycles in the Loess Plateau of
[ 66 ] Sayer E J,Powers J S,Tanner E V. Increased litterfall in China[J] . Scientific Reports ,2015 ,5: 13826.
tropical forests boosts the transfer of soil CO2 to the [ 79 ] Zhang S,Yu Z G,Lin J J,et al . Responses of soil carbon
atmosphere[J]. PLoS One,2007,2 ( 12 ) :e1299 . https: decomposition to drying-rewetting cycles : A
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]() //doi .org/ 10 . 1371/journal .pone .0001299 .
//doi .org/ 10 . 1371/journal .pone .0001299 . ![]()
![]()
![]() meta-analysis [J] .
meta-analysis [J] .![]() Geoderma,
Geoderma,![]()
![]() 2020 ,361:
2020 ,361:![]() 114069.
114069.![]()
1392
土 壤 学 报
58 卷
[ 80 ] Stark J M ,Firestone M K. Mechanisms for soil moisture
effects on activity of nitrifying bacteria[J]. Applied and
Environmental Microbiology ,1995,61 ( 1 ):218—221 .
[ 81 ] Denef K , Six J ,Bossuyt H ,et al . Influence of dry-wet
cycles on the interrelationship between aggregate ,
particulate organic matter , and microbial community
dynamics[J] . Soil Biology and Biochemistry ,2001 ,33
( 12/13 ): 1599— 1611 .
[ 82 ] Fierer N , Schimel J P , Holden P A . Influence of
drying-rewetting frequency on soil bacterial community
structure[J] . Microbial Ecology,2003,45 ( 1 ):63—71 .
[ 83 ] Magid J ,Kjærgaard C ,Gorissen A ,et al. Drying and
rewetting of a loamy sand soil did not increase the
turnover of native organic matter , but retarded the
decomposition of added 14 C-labelled plant material[J].
Soil Biology and Biochemistry,1999,31 ( 4 ):595—602 .