
生育期干旱-复水对夏玉米生化指标的影响
![]() 节水灌溉·2021 年第 6 期 jsgu.cbpt.cnki.net 9
节水灌溉·2021 年第 6 期 jsgu.cbpt.cnki.net 9
文章编号:1007-4929(2021)06-0009-08
生育期干旱-复水对夏玉米生化指标的影响
赵 龙1,2,3,蔡焕杰1,2,3,曹玉鑫1,2,3
(1. 西北农林科技大学水利与建筑工程学院,陕西 杨凌 712100;2. 西北农林科技大学中国旱区农业节水研究院,陕西 杨凌 712100;3. 西北农林科技大学旱区农业水土工程教育部重点实验室,陕西 杨凌 712100)
摘 要:以玉米品种“吉祥1 号”为试验材料,通过桶栽试验,在玉米苗期(M)和拔节期(B)进行不同程度[轻度干旱胁迫(W1)、中度干旱胁迫(W2)、重度干旱胁迫(W3)]以及不同时长[5 d(T1)、10 d(T2)、15 d(T3)]的干旱-复水处理,探究夏玉米干旱-复水过程中生化指标的变化规律和夏玉米保护酶系统的响应机制,进而探究夏玉米对干旱- 复水的响应机理。对夏玉米生育期5 个生化指标(MDA、SOD、POD、CAT 与GSH)的变化响应进行比较与分析,结果表明拔节期的各指标峰值(活性、含量)均高于对应的苗期峰值,胁迫时长为10 d 的中度胁迫处理组各指标峰值(活性、含量)均高于其他处理组。因此夏玉米苗期抗旱能力不强,不宜开展持续时间较久或干旱程度较重的亏水灌溉 方案;拔节期适宜采用持续时间为10 d 灌水量为55%~65% 土壤田间持水量的胁迫处理方案进行灌溉。
关键词:夏玉米;生育期;水分胁迫;干旱-复水;胁迫时长;生化指标;亏水灌溉;调亏灌溉;控制灌溉; 水分亏缺
中图分类号:S274.3 文献标识码:A
赵 龙,蔡焕杰,曹玉鑫. 生育期干旱-复水对夏玉米生化指标的影响[J]. 节水灌溉,2021(6):9-16.
ZHAO L,CAI H J,CAO Y X. Effect of drought stress and re-watering on biochemical indexes of summer maize during growth period[J].
Water Saving Irrigation,2021(6):9-16.
Effect of Drought Stress and Re-watering on Biochemical Indexes of Summer Maize During Growth Period
ZHAO Long1,2,3,CAI Huan-jie1,2,3,CAO Yu-xin1,2,3
(1.School of Water Conservancy and Architectural Engineering,Northwest A&F University,Yangling 712100,Shaanxi Province,China;2.Agricultural Water Conservation Research Institute of Arid Zone,Northwest A & F University,Yangling 712100,Shaanxi Province,China;3.Key Laboratory of Agricultural Soil and Water Engineering in Arid and Semiarid Areas, Ministry of Education,Yangling 712100,Shaanxi Province,China)
Abstract:The corn variety Jixiang 1 was used as the test material in this study. Through the barrel planting experiment,at the corn seedling stage (M) and jointing stage (B),the drought stress and re-watering treatment were carried out with different drought stress degrees, including mild drought stress W1,moderate drought stress W2 and severe drought stress W3,and different durations,including 5 days T1, 10 days T2 and 15 days T3,to explore the change rule of biochemical indexes and the response mechanism of summer maize protective enzyme system in the process of drought and re-watering,and then to explore the response mechanism of summer maize to drought and re- watering. The responses of five biochemical indexes (MDA,SOD,POD,CAT and GSH) in summer maize growth stages were compared and analyzed. The results showed that the peak values (activity and content) of each index at jointing stage were higher than those at seedling stage,and the peak values (activity and content) of each index in moderate stress group were higher than those in other treatment groups under stress duration of 10 days. Therefore,the drought resistance of summer maize seedling stage is not strong,so it is not suitable to carry out the irrigation scheme of deficit water with longer duration or more severe drought degree. At jointing stage,it is suitable to adopt the stress treatment scheme with the irrigation amount of 55%~65% soil field capacity for 10 days.
![]()
Key words:summer maize;growth stages;water stress;drought stress and re-watering;stress duration;biochemical indicators;deficit water irrigation;regulated deficit irrigation;controlled irrigation;water deficit
收稿日期:2021-01-18
基金项目:国家自然科学基金项目(51879223);国家重点研发计划项目(2016YFC0400201)。
作者简介:赵 龙(1996-),男,硕士研究生,主要研究方向为灌溉排水新技术。E-mail:854296432@qq.com。
通讯作者:蔡焕杰(1962-),男,教授,主要从事农业节水与水资源高效利用研究。E-mail: caihj@nwsuaf.edu.cn。
![]()
![]() 10
10
0 引 言
生育期干旱-复水对夏玉米生化指标的影响 赵 龙 蔡焕杰 曹玉鑫
胱甘肽含量。
1.4 试验方法
![]()
全球变暖加剧了目前干旱发生的频率和程度,粮食生产
已经受到了严重威胁[1]。已有研究表明,水分胁迫会抑制作物 的生长,降低作物产量[2]。玉米是需水量大、对水分胁迫敏感
本试验研究将桶栽夏玉米的其中2 个生长阶段进行试验处理,分别是夏玉米的苗期(M)和拔节期(B)。设有3 个水分处理, 其中,对照组(CK):75%~85% θ (θ 表示为田间持水量);轻度
的粮食作物,然而干旱导致玉米每年减产20%~30% [3, 4]。中国 f f
西北部地处内陆, 气候干燥, 有大面积的干旱和半干旱地区[5]。玉米是我国西北地区的主要粮食作物之一,该地区玉米 产量也一直受限于水资源匮乏问题[4, 6, 7]。因此,培育高抗性玉米品种是应对干旱胁迫最有效,也是最重要的途径[8]。所以了 解玉米对干旱胁迫的生理响应机制是抗旱育种的重要前提[9-11]。
干旱胁迫对作物的生成会产生一定的危害,然而干旱后恢复至正常水平灌溉后会使植物产生补偿甚至超补偿效应, 并且大量研究证明,适度的干旱胁迫后复水作物所产生的补偿效应可以使作物的产量或品质得以提高[12]。干旱胁迫打破了植物体内活性氧产生和清除之间的平衡[13],为了清除因外界胁迫大量累积的活性氧,植物体内的抗氧化酶活性增加,抗氧化物质合成增多[14],从而减轻胁迫对细胞膜造成的伤害,起到对植物的保护作用[15]。这种保护可以有效减小细胞的氧化损
伤,使得恢复正常灌溉水平后植物能够迅速得以恢复[16]。
干旱胁迫(W1):65%~75% θf ;中度干旱胁迫(W2):55%~65% θf ;重度干旱胁迫(W3):45%~55% θf 。并设置3 个胁迫时长处理,分别是5 d(T1)、10 d(T2)、15 d(T3),具体处理方案见表1。控水方法为称重法。共计16 个处理,CK 对照处理组设置5 个重复,苗期处理设置4 个重复,拔节期处理设置3 个重复,共 计种植夏玉米 59 桶。由于CK 对照组在整个试验期间均需取样,苗期夏玉米的叶片数量较少,故多设置了试验重复桶。 试验期间于胁迫期间每 5 d 取 3 次样,复水后 10 d 内再取 3 次样,均在浇水前取样;其余生育期灌水制度均与CK 对照处理相同。试验处理期间每天称一次桶重,称重后进行浇水,时 间为每天下午6∶00。
表 1 试验设计
Tab.1 Experimental design
![]()
已有对作物干旱-复水这一过程的研究主要集中于不同灌溉水平对作物产量和水分利用效率的影响,本研究对夏玉米
苗期(M)
T1
![]() T2 CK
T2 CK
MW1T1 MW1T2
MW2T1 MW2T2
MW3T1 MW3T2
苗期和拔节期进行不同灌水水平和不同胁迫时长的干旱-复水
T3 MW1T3 MW2T3 MW3T3
处理, 通过 5 个生化指标[ 丙二醛(MDA)、超氧化物歧化酶
(SOD)、过氧化物酶(POD)、过氧化氢酶(CAT)、还原型谷胱甘
T1
拔节期(B) T2
BW1T1
CK BW1T2
BW2T1 BW2T2
肽(GSH)]的变化响应进行研究分析,探究夏玉米保护酶系统对 干旱-复水处理的响应机制,从而找出对夏玉米产量较有利的 干旱胁迫方案,以期对夏玉米的高效节水灌溉提供理论支持。
1 材料与方法
1.1 材 料
试验对象为夏玉米,所用品种“吉祥1 号”是当地农民普遍使用的品种,该品种根系发达,叶片多且上举。本次试验 玉米生长期为2019 年6 月16 日至2019 年9 月22 日,共计98 d。桶栽种植玉米用土的土壤类型为中壤土,沙和黏粒含量占比 大致相同,具有一定可塑性,土壤偏弱碱性。田间持水量为24%(重量含水率),土壤干容重为 1.33 g/cm3。按 200 kg/hm2 的施氮量、100 kg/hm2 的施磷量,换算出每桶施基肥尿素2.37 g, 磷酸二铵4.75 g[17],于2019 年8 月4 日追肥尿素 3 g。试验用桶栽的方式种植玉米,采用30 cm×30 cm×40 cm(0.036 m3)的试验方桶。
1.2 试验区概况
试验地点地处东经 108°04',北纬 34°18',海拔 521 m 的西北农林科技大学中国旱区节水农业研究院,所有试验方桶 均放置于遮雨棚下。
1.3 指标测定
丙二醛含量用硫代巴比妥酸反应比色法测定;NBT 光化学还原法测定超氧化物歧化酶活性;愈创木酚法测定过氧化 物酶活性;紫外分光光度法测定过氧化氢酶活性和还原性谷
T3 BW1T3 BW2T3
1.5 数据统计
采用 Origin 进行绘图,采用 SPSS 26.0 对试验数据进行统计分析。
2 结果与分析
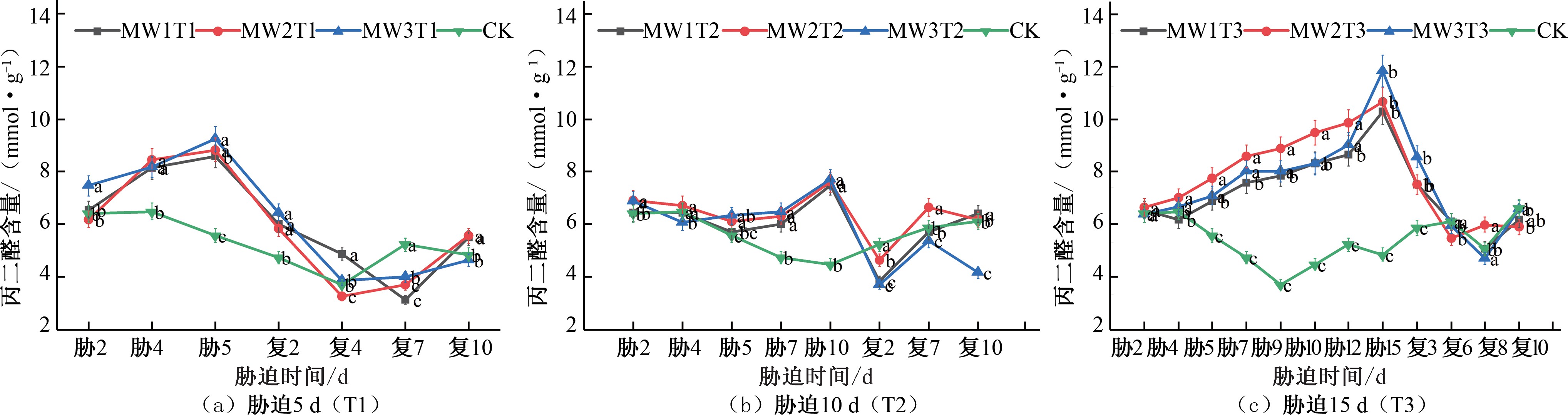
2.1 干旱胁迫-复水对夏玉米丙二醛(MDA)含量的影响
植物在受到外界环境胁迫时,抵抗胁迫的过程中会生成丙二醛(MDA),植物细胞膜系统在经受胁迫时会有过氧化反 应,而MDA 含量的高低可以判断过氧化的程度,因此它可以反应植物在胁迫条件下损伤的程度[18]。由图1 可知,在夏玉米苗期期间,各试验组在进行胁迫处理期间,随着胁迫时长的增长,干旱程度由轻度到重度的加剧,MDA 累积量逐渐增多, 每组试验均在胁迫的最后一天均达到峰值。MW1T1、MW2T1 和MW3T1 处理在胁迫第 5 天MDA 含量分别为CK 对照处理的
1.54 倍、 1.58 倍、 1.66 倍[ 见图 1(a)]; MW1T2、 MW2T2 和
MW3T2 处理在胁迫第 10 天 MDA 含量分别为CK 对照处理的
1.67 倍、 1.69 倍和 1.71 倍[ 见图 1(b)]; MW1T3、 MW2T3 和
MW3T3 处理在胁迫第 15 天MDA 含量分别为CK 对照处理的
2.12 倍、2.20 倍和 2.43 倍[见图 1(c)]。在进行复水后,胁迫程度得到缓解, MDA 含量立即降低趋向于CK 对照处理水平, MW1T1、MW2T1 和MW3T1 处理在复水后第7 天出现明显补偿效应,MDA 含量分别为CK 对照处理的0.60 倍、0.71 倍和0.76
![]()
生育期干旱-复水对夏玉米生化指标的影响 赵 龙 蔡焕杰 曹玉鑫 11
![]()
![]()
![]()
![]()
倍; MW1T2 和MW2T2 处理在复水第 2 天出现了补偿效应, MDA 含量分别为CK 对照处理的 0.73 倍和 0.88 倍,MW3T2 处理复水 2 d 后出现了补偿效应,复水第 10 天补偿效应最为明显,MDA 含量为CK 对照处理的0.68 倍;胁迫时长为15 d 的处理组复水后未出现明显补偿效应。在水分胁迫处理期间,各
试验组的MDA 含量随胁迫时长增长均呈递增趋势,中度水分胁迫和重度水分胁迫的MDA 含量峰值相近且均高于轻度水分胁迫,说明干旱胁迫程度的不断加重,夏玉米苗期细胞内的 活性氧含量也随之增大,细胞膜过氧化作用加强,MDA 累积量也随之增多。
八1_∞.一oIIIIII
图1 夏玉米苗期丙二醛(MDA)含量于干旱胁迫-复水处理下的变化
![]() Fig.1 Changes of MDA c八1_∞.o一oIIIIIIntent in summer maize seedlings under drought stress and八1_∞.一oIrIIIIIehydration treatment
Fig.1 Changes of MDA c八1_∞.o一oIIIIIIntent in summer maize seedlings under drought stress and八1_∞.一oIrIIIIIehydration treatment
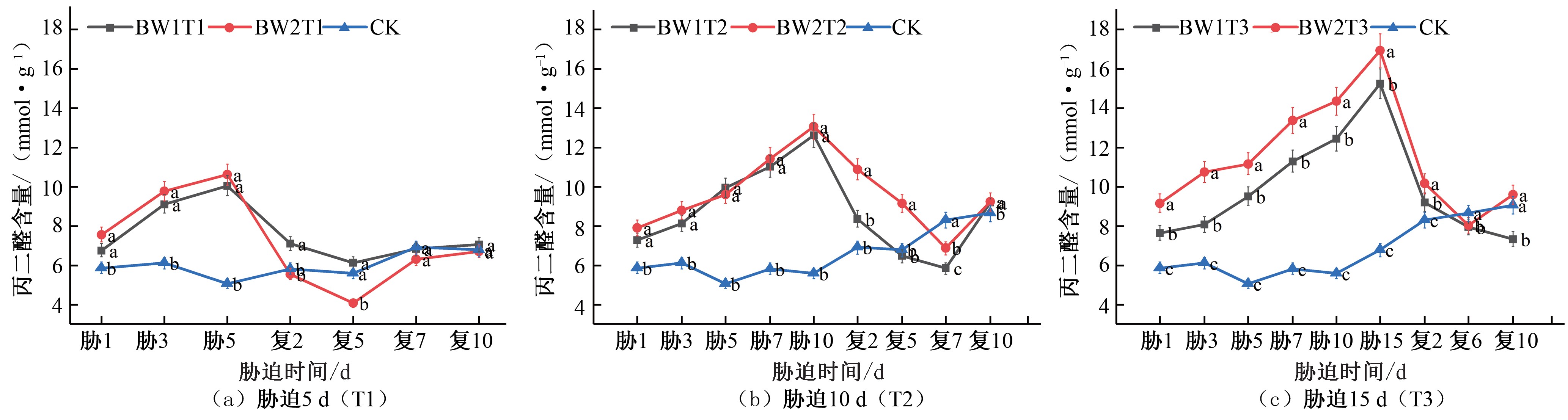 拔节期的MDA 变化趋势与苗期相同,均随着胁迫持续时间的延长,干旱胁迫程度的加剧,MDA 累积量变大,每组试验均在胁迫的最后一天均达到峰值。BW1T1 和BW2T1 处理在胁迫末期MDA 含量分别为CK 对照处理的 1.98 倍和 2.09 倍[见图 2(a)]; BW1T2 和BW2T2 处理在胁迫末期MDA 含量分别为CK 对照处理的 2.15 倍和 2.34 倍[见图 2(b)];BW1T3 和BW2T3 处理在胁迫末期MDA 含量分别为CK 对照处理的2.25 倍和2.50 倍[见图 2(c)]。在不同的水分胁迫处理下,中度水分胁迫处理相较于轻度胁迫处理,其MDA 积累量更大,细胞膜的过氧化程度更为明显。在进行了复水处理之后,各试验组MDA 含量随着胁迫程度缓解降低,趋向于CK 对照处理水平。BW1T1 处理在复水后未出现明显补偿效应,BW2T1 处理在复水后第 5 天出现了明显的补偿效应,MDA 含量为CK 对照处理的 0.73 倍;BW1T2 和BW2T2 处理在复水后第 7 天时出现补偿效应, MDA 含量分别为CK 对照处理的 0.70 倍和 0.82 倍;BW1T3 处
拔节期的MDA 变化趋势与苗期相同,均随着胁迫持续时间的延长,干旱胁迫程度的加剧,MDA 累积量变大,每组试验均在胁迫的最后一天均达到峰值。BW1T1 和BW2T1 处理在胁迫末期MDA 含量分别为CK 对照处理的 1.98 倍和 2.09 倍[见图 2(a)]; BW1T2 和BW2T2 处理在胁迫末期MDA 含量分别为CK 对照处理的 2.15 倍和 2.34 倍[见图 2(b)];BW1T3 和BW2T3 处理在胁迫末期MDA 含量分别为CK 对照处理的2.25 倍和2.50 倍[见图 2(c)]。在不同的水分胁迫处理下,中度水分胁迫处理相较于轻度胁迫处理,其MDA 积累量更大,细胞膜的过氧化程度更为明显。在进行了复水处理之后,各试验组MDA 含量随着胁迫程度缓解降低,趋向于CK 对照处理水平。BW1T1 处理在复水后未出现明显补偿效应,BW2T1 处理在复水后第 5 天出现了明显的补偿效应,MDA 含量为CK 对照处理的 0.73 倍;BW1T2 和BW2T2 处理在复水后第 7 天时出现补偿效应, MDA 含量分别为CK 对照处理的 0.70 倍和 0.82 倍;BW1T3 处
理在复水后第 10 天出现了明显的补偿效应,MDA 含量为CK 对照处理的 0.81 倍, BW2T3 处理在复水后未出现明显补偿效应。
综合夏玉米苗期和拔节期不同灌溉水平下MDA 含量的变化,结果表明作物在受到水分胁迫时,由于累积大量活性氧 自由基不能及时被清除,细胞膜的结构受到了损伤,MDA 含量也随之升高,重度水分胁迫处理的MDA 含量与中度水分胁迫相近,中度水分胁迫处理的MDA 含量高于轻度水分胁迫; 随着胁迫持续时间越长,各试验组MDA 含量越高。这与郭效龙等人的研究结论相一致[19, 20]。在夏玉米苗期MDA 的最高含量达到 11.82 mmol/g, 拔节期 MDA 的最高含量达到 16.96 mmol/g,所以夏玉米拔节期的抗旱能力高于苗期的抗旱能力。复水后胁迫缓解,所有处理组的MDA 含量均逐渐恢复至CK 对照处理。
![]() v\删如龃¨瞌
v\删如龃¨瞌
图2 夏玉米拔节期丙二醛(MDA)含量于干旱胁迫-复水处理下的变化
Fig.2 Changes of MDA content in summer maize at jointing stage under drought stress and rehydration treatment
2.2 干旱胁迫-复水对夏玉米超氧化物歧化酶(SOD) 活性的影响
植物抗氧化系统中,超氧化物歧化酶(SOD)作为第 1 道防
线,它的功能是加速超氧物阴离子自由基进行氧化还原反应,
胁迫5 d 的处理组在胁迫第4 天胁时2达胁到4胁峰5胁值7,胁相9胁较1于0胁C1K2胁对1照5复处3复6复8复10理MW1T1、MW2T1、MW3T1 的SOD 活性分别提高 76.87%、
49.58%、46.62% [见图3(a)];胁迫10 d 的处理组在胁迫第5 天达到峰值,相较于CK 对照MW1T2、MW2T2、MW3T2 的SOD
产生过氧化胁氢迫以时及间氧/d气[21]。由图3 可知,在夏玉米苗期,胁迫胁迫时活间性/处d理分别提高13.19%、26.33%、48.45%[见胁迫图时3(间b)/];d胁迫
期间所有试验组的SOD 活性的变化趋势均为先增大后减小,
15 d 的处理组在胁迫第 7 天达到峰值, 相较于CK 对照处理
6
![]()
12 生育期干旱-复水对夏玉米生化指标的影响 赵 龙 蔡焕杰 曹玉鑫
![]()
![]()
![]()
![]()
4
2
八 T_I: 【 一 III.T_M.≥v O
MW1T3、 MW2T3、 MW3T3 的 SOD 活性分别提高 25.50%、
30.95%、33.96%[ 见图 3(c)]。各试验组均在胁迫处理的末期SOD 活性降到最低点,复水处理后,SOD 活性先增大后减小, 各试验组均出现了一定的补偿效应,胁迫5 d 的试验组均于复水第 2 天SOD 活性达到峰值, 相较于CK 对照处理MW1T1、MW2T1、 MW3T1 的 SOD 活 性 分 别 提 高 7.38%、 14.87%、
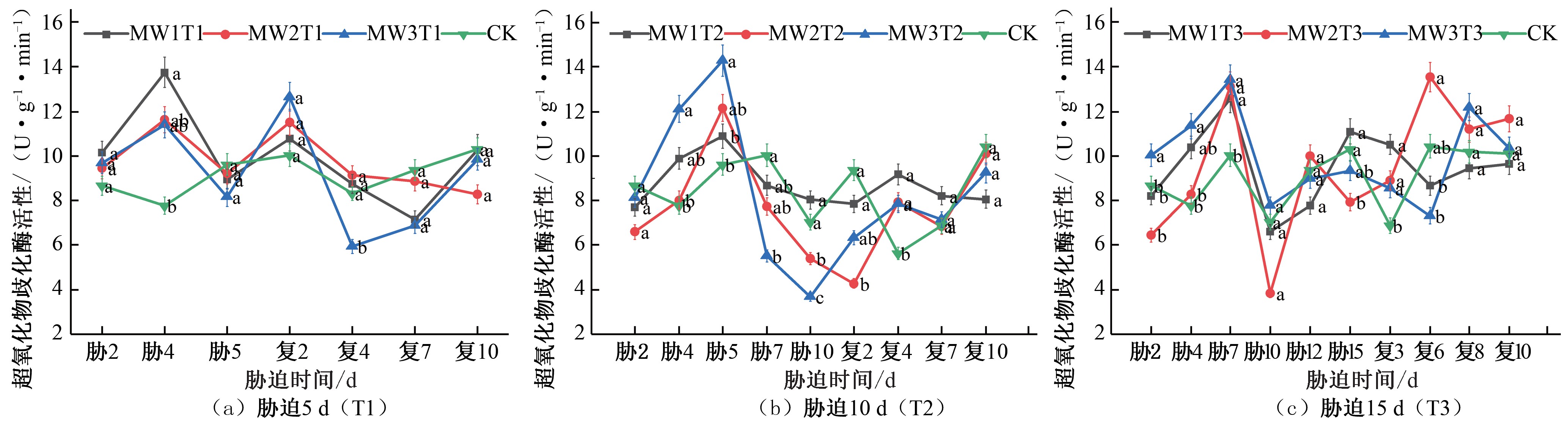 26.25%[见图 3(a)];胁迫 10 d 的试验组均于复水第 4 天SOD 活性达到峰值, 相较于CK 对照MW1T2、MW2T2、MW3T2 的SOD 活性处理分别提高 63.06%、41.02%、39.51%[见图 3(b)];
26.25%[见图 3(a)];胁迫 10 d 的试验组均于复水第 4 天SOD 活性达到峰值, 相较于CK 对照MW1T2、MW2T2、MW3T2 的SOD 活性处理分别提高 63.06%、41.02%、39.51%[见图 3(b)];
胁迫15 d 的试验组MW1T3 于复水第3 天达到峰值,MW2T3 于复水第6 天达到峰值,MW3T3 于复水第8 天达到峰值,相较于CK 对照处理MW1T3、MW2T3、MW3T3 的SOD 活性分别提高52.71%、29.70%、19.44%[见图 3(c)]。达到峰值后各试验处理组SOD 活性呈下降趋势,最终变化趋势趋向于CK 对照处理水平。综合3 个不同胁迫时长处理的结果,可以看出胁迫时长为10 d 的处理组在复水后SOD 活性相较于CK 对照处理增大得最多,出现的补偿效应最为明显。
图3 夏玉米苗期超氧化物歧化酶(SOD)活性于干旱胁迫-复水处理下的变化
Fig.3 Changes of SOD activity in summer maize seedlings under drought stress and rehydration treatment
夏玉米拔节期期间,胁迫5 d 处理组的夏玉米,在胁迫期间SOD 活性均呈现增大的趋势,BW1T1 在胁迫第 5 天SOD 活性达到最高点,BW2T1 于胁迫第3 天达到峰值,相较于CK 对照处理分别提高了 5.91% 和 14.36%;复水后,试验组SOD 活性均呈下降趋势[见图 4(a)]。胁迫 10 d 处理组的夏玉米,在胁迫期间BW1T2 的SOD 活性呈现先增大后减小的趋势,于胁迫第5 天达到峰值,相较于CK 对照处理提高了34.87%,BW2T2 的SOD 活性始终低于CK 对照处理水平;复水后胁迫程度缓解,复水第 2 天SOD 活性降至最低,之后BW1T2 的SOD 活性先增大后减小,在复水第 5 天SOD 活性达到最高点,相较于CK 对照处理提高了51.98%,出现明显补偿效应,BW2T2 的则
逐渐增加恢复至CK 水平[见图 4(b)]。胁迫 15 d 处理组的夏玉米,在胁迫期间SOD 活性均呈现先增大后减小的趋势,于胁迫第5 天达到峰值,BW1T3 和BW2T3 的SOD 活性相较于CK 对照处理分别提高了 22.92% 和 18.57%;复水后BW1T3 变化趋势与CK 对照处理组相一致,BW2T3 在复水后第2 天SOD 活性相较于CK 对照处理提高了54.67%,之后降低至CK 水平[见图4(c)]。在夏玉米拔节期,所有处理均在胁迫第 5 天达到峰值, 说明在拔节期胁迫第5 天时,作物所遭受的胁迫状态相同。复水后,胁迫时长为10 d(T2)处理组补偿效应较为明显,其SOD 活性明显高于胁迫时长为5 d(T1)和15 d(T3)的处理组。
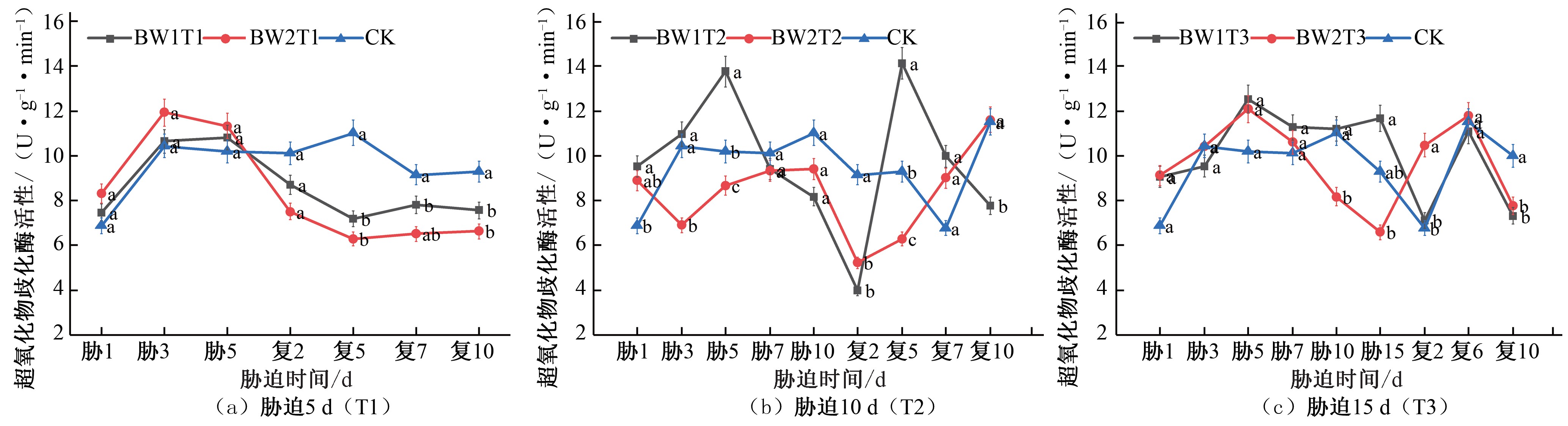
图4 夏玉米拔节期超氧化物歧化酶(SOD)活性于干旱胁迫-复水处理下的变化
Fig.4 Changes of SOD activity in summer maize at jointing stage under drought stress and rehydration treatment
![]() 综合夏玉米苗期和拔节期在干旱胁迫-复水过程中SOD 活性的变化响应,相较于CK 对照处理,复水后SOD 活性呈现先增大后减小的趋势,这与庄克章的研究结论相一致[22]。胁迫时长为 10 d 的处理组复水后补偿效应最明显,不同胁迫程度的
综合夏玉米苗期和拔节期在干旱胁迫-复水过程中SOD 活性的变化响应,相较于CK 对照处理,复水后SOD 活性呈现先增大后减小的趋势,这与庄克章的研究结论相一致[22]。胁迫时长为 10 d 的处理组复水后补偿效应最明显,不同胁迫程度的
2.3 干旱胁迫-复水对过氧化物酶(POD)活性的影响
![]() 过氧化物酶(POD)是植物体细胞内普遍存在的,植物的代 谢和抗逆性都与其有着紧密的关系[23],它的主要作用是还原过氧化氢,将其转变成水分子,通过这个过程来清除过氧化氢。
过氧化物酶(POD)是植物体细胞内普遍存在的,植物的代 谢和抗逆性都与其有着紧密的关系[23],它的主要作用是还原过氧化氢,将其转变成水分子,通过这个过程来清除过氧化氢。
\赳,蜉瀣睾督S睾噼裂
处理组的对胁比迫中时间可/以d看出中度胁迫处理八下口IⅢ.1∞.≥v\赳,的蜉瀣睾督S睾噼裂
SOD 活性复水后高
在夏玉米苗期,胁迫时长八下口IⅢ.1∞.≥v\赳,蜉瀣睾督S睾噼裂为5 d 的试验组MW1T1 和MW2T1 在
胁迫期间POD 活性不断增大,并在胁迫第5 天达到峰值,分别
![]()
生育期干旱-复水对夏玉米生化指标的影响 赵 龙 蔡焕杰 曹玉鑫 13
![]()
![]()
![]()
![]()
相对于CK 对照组增加了24.88% 和55.28%,MW3T1 的POD 活性先减小后增大,胁迫期间低于CK 对照处理组水平;复水处理后,各试验组均在复水第4 天达到峰值,MW1T1、MW2T1、MW3T1 的 POD 活性分别相对于 CK 对照组增加了 32.97%、16.53%、29.70%,出现了一定补偿效应,之后降低至CK 对照处理水平[ 见图 5(a)]。胁迫时长为 10 d 的试验组 MW2T2 和MW3T2 分别在胁迫第 5 天和第 9 天POD 的活性达到峰值,分别相对于CK 对照组增加了37.66% 和94.94%,MW1T2 的试验组在胁迫期间POD 活性始终低于CK 对照处理水平;复水处理
后,各处理的POD 活性不断增加,MW1T2 和MW2T2 的POD 活性复水后始终低于CK 对照处理水平,MW3T2 的POD 活性在复水第7 天后高于CK 对照处理,出现一定补偿效应[见图5 (b)]。胁迫时长为15 d 的试验组POD 活性均呈现先增大后减小的趋势,MW1T3 和MW3T3 在胁迫第 5 天的POD 活性达到峰值,分别相对于CK 对照组增加了 31.01% 和 28.22%,MW2T3 在胁迫第9 天的POD 活性达到峰值,相对于CK 对照组增加了18.02%;复水后POD 活性有所增加,但均未出现明显补偿效应[见图5(c)]。
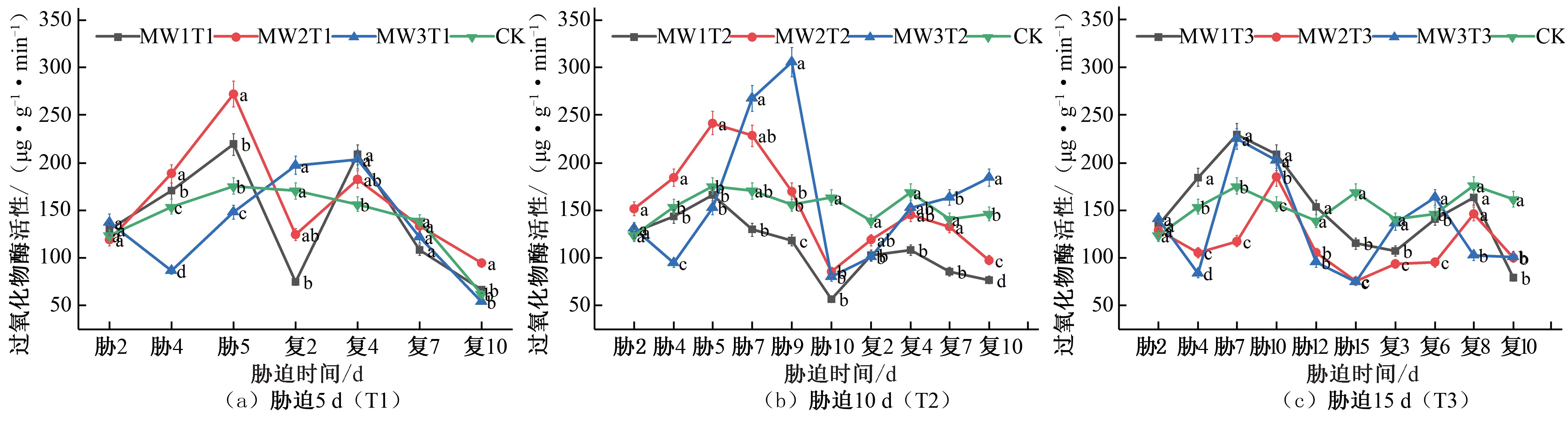
图5 夏玉米苗期过氧化物酶(POD)活性于干旱胁迫-复水处理下的变化
Fig.5 Changes of POD activity in summer maize seedlings under drought stress and rehydration treatment
在夏玉米拔节期,胁迫5 d 的试验组POD 活性在胁迫期间BW1T1 持续增大,于胁迫第5 天达到峰值,相较于CK 对照组提升了46.14%,复水后先减小后增大,逐渐增大至CK 对照水平;BW2T1 的POD 活性在夏玉米拔节期始终低于CK 对照水平[见图6(a)]。胁迫10 d 的处理组在胁迫期间BW1T2 在胁迫第10 天出现峰值,相较于CK 对照组提升了 39.44%,BW2T2 胁迫
期间POD 活性始终低于CK 对照水平,复水后两组处理POD 活性均显著增加,均在复水后第7 天POD 活性达到峰值,出现补偿效应,分别相较于CK 对照组提升了55.19%、13.85%,之后降低至CK 对照水平[见图6(b)]。胁迫15 d 的处理组在胁迫期间POD 活性均低于CK 对照水平,复水后虽然有所增加,但始终低于对照组水平[见图6(c)]。
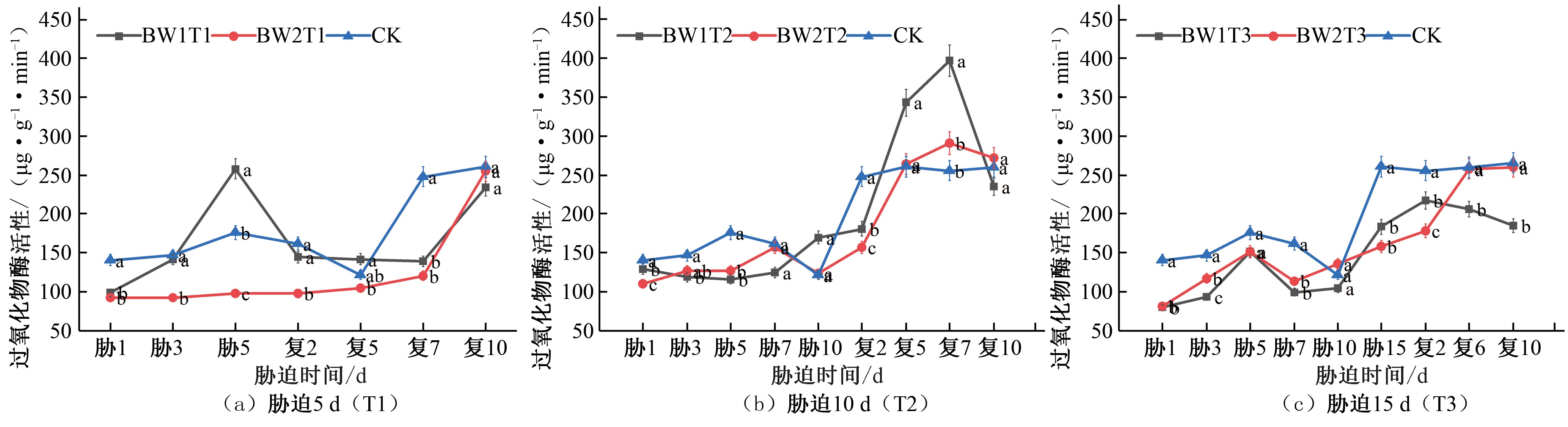
图6 夏玉米拔节期过氧化物酶(POD)活性于干旱胁迫-复水处理下的变化
Fig. 6 Changes of POD activity in summer maize at jointing stage under drought stress and rehydration treatment
综合对比夏玉米苗期和拔节期在干旱胁迫-复水过程中POD 活性的变化响应,随着胁迫时长的增长,POD 活性相较于CK 对照处理增大幅度逐渐降低,说明由于干旱胁迫程度不断加剧,植物体内抗氧化系统的工作出现了失调。恢复正常 水平灌水后各处理的POD 活性均有所增加,这与黄旭光研究朱槿幼苗的POD 活性变化规律一致[24]。拔节期的POD 活性明
的转移而最终将其分解为水和氧气[25],避免了过氧化氢在体内的积累,从而可维持体内正常的活性氧水平[26]。由图 7 可知, 在夏玉米苗期期间,胁迫5 d 的处理组在胁迫期间CAT 活性先增大后减小, 在胁迫第 4 天达到峰值, MW1T1、MW2T1 和MW3T1 的 CAT 活性相较于 CK 对照组分别提升了 64.49%、104.23%、52.18%,而第5 天减小至最小值;复水后所有试验
![]() 八1_口IⅢ.1∞.望v\赳,蜉瀣S睾噼蜊
八1_口IⅢ.1∞.望v\赳,蜉瀣S睾噼蜊
显高于苗期, 说明夏玉米拔节期清除八下口IⅢ.1过∞.望v\赳,蜉瀣S睾噼蜊
氧化氢的能力强于
组CAT 活性先升高后降低,MW1T1 水平于复水第 2 天达到峰值,MW2T1 和MW3T1 于复水第 4 天达到峰值,相较于CK 对
![]()
![]()
![]() 2.4 干旱胁迫-复水对夏玉米过氧化氢酶(CAT)活性的影响
2.4 干旱胁迫-复水对夏玉米过氧化氢酶(CAT)活性的影响
过氧化氢酶(CAT)以过氧化氢为底物,通过催化一对电子
照组分别提升了 11.19%、79.37%、32.53%,最终 3 个水分胁迫处理组的CAT 活性均降低至CK 对照处理水平[见图7(a)]。胁迫10 d 的3 组不同的水分处理胁迫期间均在胁迫第4 天CAT 活
![]()
14 生育期干旱-复水对夏玉米生化指标的影响 赵 龙 蔡焕杰 曹玉鑫
![]()
![]()
![]()
![]()
性达到峰值,MW1T2、MW2T2 和MW3T2 的CAT 活性相较于CK 对照组分别提升了31.16%、55.07%、36.73%,而第5 天减小至最小值,在胁迫末期均低于CK 对照处理水平;复水后, MW1T2 和MW2T2 均先增大后减小,MW1T2 的CAT 活性在复水后第 2 天达到峰值,MW2T2 的CAT 活性在复水后第 4 天达到峰值,分别相较于CK 对照组分别提升了 5.23%、32.43%, 出现补偿效应并最终降低至CK 水平,MW3T2 复水后呈现先减小后增大的趋势,在复水第 7 天出现补偿效应[见图 7(b)]。胁
迫15 d 的处理组,在胁迫期间均于胁迫第4 天CAT 活性达到峰值,MW1T3、MW2T3 和MW3T3 的CAT 活性相较于CK 对照组分别提升了56.38%、31.84%、26.50%,在胁迫末期CAT 活性和CK 对照处理水平一致,在复水期间3 个试验组的CAT 活性均先增大后减小,出现补偿效应,MW1T3 和MW2T3 在复水第 3 天达到峰值, 相较于 CK 对照组分别提升了 18.58%、
27.07%,MW3T3 在复水第8 天达到峰值,相较于CK 对照组提升了40.28%,之后均降低至CK 水平[见图7(c)]。
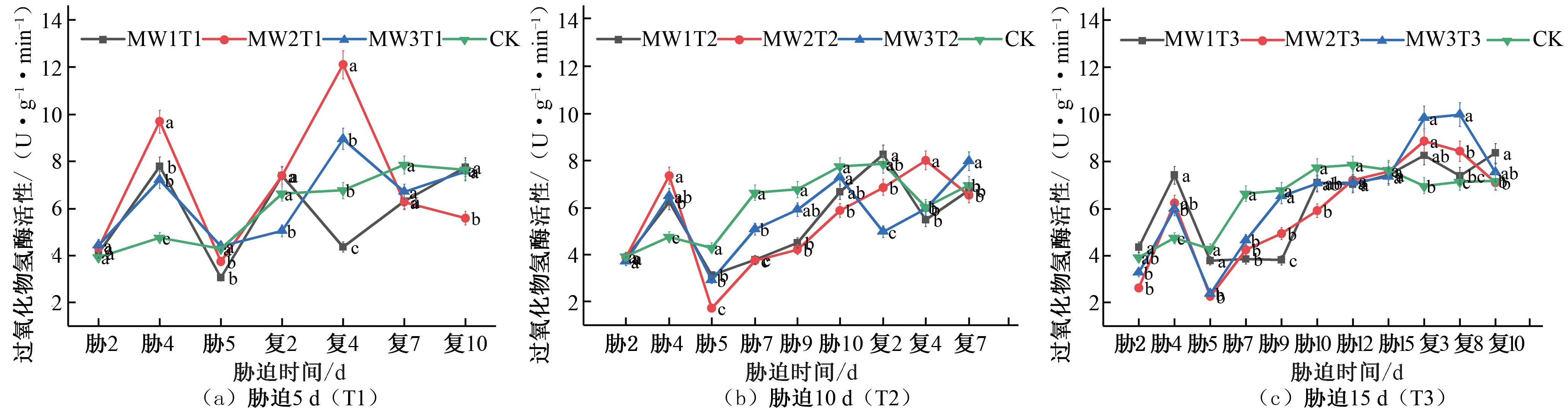
图7 夏玉米苗期过氧化氢酶(CAT)活性于干旱胁迫-复水处理下的变化
Fig.7 Changes of CAT activity in summer maize seedlings under drought stress and rehydration treatment
在夏玉米拔节期期间,胁迫5 d 的处理组,BW1T1 的CAT 活性在胁迫期间先增大后减小,在胁迫第3 天达到峰值,相较于CK 对照组提升了52.94%,复水后也呈现先增加后减小的趋势, 于复水第 5 天达到峰值, 相较于 CK 对照组提升了53.31%;BW2T2 的CAT 活性在胁迫期间持续增加至胁迫第 5 天达到峰值,相较于CK 对照组提升了32.55%,复水后先减小后增加,于复水后第7 天达到峰值,相较于CK 对照组提升了38.13%,2 个不同水分水平的处理均出现补偿效应且于复水后第10 天恢复至CK 水平[见图8(a)]。胁迫10 d 的2 个水分处理组CAT 活性均在胁迫期间先升高后降低,胁迫第5 天达到峰值, BW1T2 和 BW2T2 相较于 CK 对照组分别提升了 74.23%、
38.54%;复水后各试验组也均呈现先增大后减小的趋势,并 于复水后第2 天达到峰值,CAT 活性相较于CK 对照组分别提升了 5.93%、38.07%,出现补偿效应,之后CAT 活性开始减小,复水后第7 天降低至CK 水平[见图8(b)]。胁迫15 d 的2 个处理组在胁迫期间先升高后降低,在胁迫第3 天的时候达到峰值,BW1T3 和BW2T3 的CAT 活性相较于CK 对照组分别提升了 33.44%、16.15%; 复水后CAT 活性持续增加, BW1T3 和BW2T3 出现明显补偿效应,其CAT 活性分别于复水后第10 天和复水后第 6 天达到峰值, 相较于 CK 对照组分别提升了35.37%、29.96%[见图8(c)]。
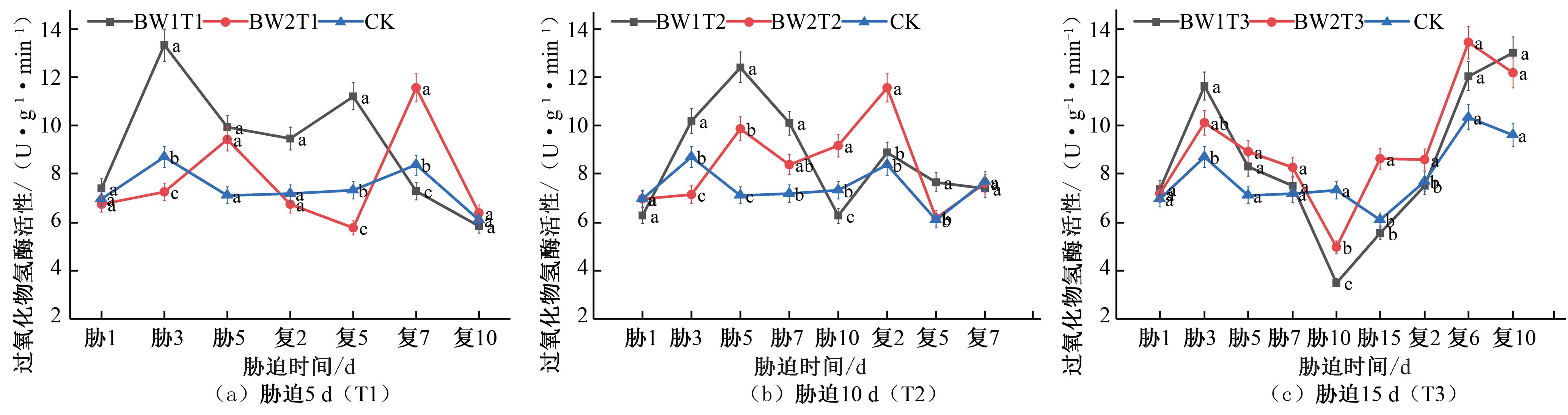
图8 夏玉米拔节期过氧化氢酶(CAT)活性于干旱胁迫-复水处理下的变化
Fig.8 Changes of CAT activity in summer maize at jointing stage under drought stress and rehydration treatment
综合比较夏玉米苗期和拔节期在干旱胁迫-复水过程中
CAT 活性的变化响应,在夏玉米苗期胁迫期间,CAT 活性均
期处理,说明作物拔节期抵抗胁迫危害的能力强于苗期。
2.5 干旱胁迫-复水对夏玉米还原型谷胱甘肽(GSH)
于胁迫第4 天达到峰值,这与刘承[27]利用PEG-6000 模拟水分
含量的影响
胁2胁4胁5胁7胁9胁10胁12胁15复3复8复10
八下口IⅢ.1h.≥v\赳,蜉瀣蛹S睾噼蜊
胁迫的研究得出的结论一致;复水后轻八下口IⅢ.1h.≥v\赳,蜉瀣蛹度S睾噼蜊
和中度水分胁迫处
植物对于外界环境胁八下口IⅢ.1h.≥v\赳,蜉瀣蛹S睾噼蜊迫的耐受性与植物体内谷胱甘肽含
理均在复水4 天内达到峰值,而重度水分胁迫处理均在复水4 天后达到峰值,说明随着胁迫程度增加,复水后CAT 响应时
量密切相关,在植物受到干旱胁迫时,植物细胞自身代谢过 程中会生成部分过氧化物,还原型谷胱甘肽(GSH)可以非常高
间增长。胁夏迫玉时米间拔/节d 期的CAT 活性均高于相同胁迫程度的苗胁迫时间效/地d清除这些过氧化物[28]。GSH 可以由植物体细胁胞迫内时的间氧/化d 型
![]()
生育期干旱-复水对夏玉米生化指标的影响 赵 龙 蔡焕杰 曹玉鑫 15
![]()
![]()
![]()
![]()
∞ 谷胱甘肽(GSSG)转化而来,GSH 含量增∞高后植物抗逆能力也随之变强[29]。在夏玉米苗期, 胁迫 5 d 的处理组 MW1T1 和
GSH 含量达到峰值,分别∞为对照组的 1.69 倍和 1.95 倍,出现明显补偿效应,MW3T2 复水后未出现补偿效应[见图9(b)]。胁
● MW2T1 的 GSH 含量呈现先减小后增大的● 趋势,在胁迫第5 天
迫15 d 的处理组MW1T3 的● GSH 含量先减少后增加再下降,于
![]()
![]() 达到峰值,分别是CK 对照处理组的1.35 倍、1.23 倍,MW3T1 的 GSH 含量胁迫期间持续降低;复水后,各试验组GSH 含量均在复水第 2 天达到峰值,分别是CK 对照处理组的 1.31 倍、
达到峰值,分别是CK 对照处理组的1.35 倍、1.23 倍,MW3T1 的 GSH 含量胁迫期间持续降低;复水后,各试验组GSH 含量均在复水第 2 天达到峰值,分别是CK 对照处理组的 1.31 倍、
i 1.16 倍、1.82 倍,出现明显补偿效应,之i后降低至CK 对照处
胁迫第7 天达到峰值,是CK 对照处理组的1.73 倍,MW2T3 和
![]() MW3T3 的GSH 含量均先增∞ 加再减少,于胁迫第4 天达到峰值, 分别是CK 对照处理组的1.73 倍和1.96 倍;在复水后3 组不同
MW3T3 的GSH 含量均先增∞ 加再减少,于胁迫第4 天达到峰值, 分别是CK 对照处理组的1.73 倍和1.96 倍;在复水后3 组不同
水分胁迫处理GSH 含量均有所增加,MW1T3 和MW2T3 分别
\/ 理水平[ 见图 9(a)]。胁迫 10 d 的处理组MW1T2 和MW2T2 的
于复水后第 8 天和第 6 天\达/ 到峰值,相较于CK 对照组GSH 含
GSH 含量在胁迫第 7 天达到峰值, 分别\/是 CK 对照处理组的
1.22 倍和1.33 倍,MW3T2 的GSH 含量胁迫期间持续降低;复水后,MW1T2 和MW2T2 分别于复水后第4 天和复水后第2 天
量分别是其1.21 倍和1.26 倍,出现了补偿效应,MW3T3 复水后GSH 含量逐渐增大,但未出现补偿效应[见图9(c)]。
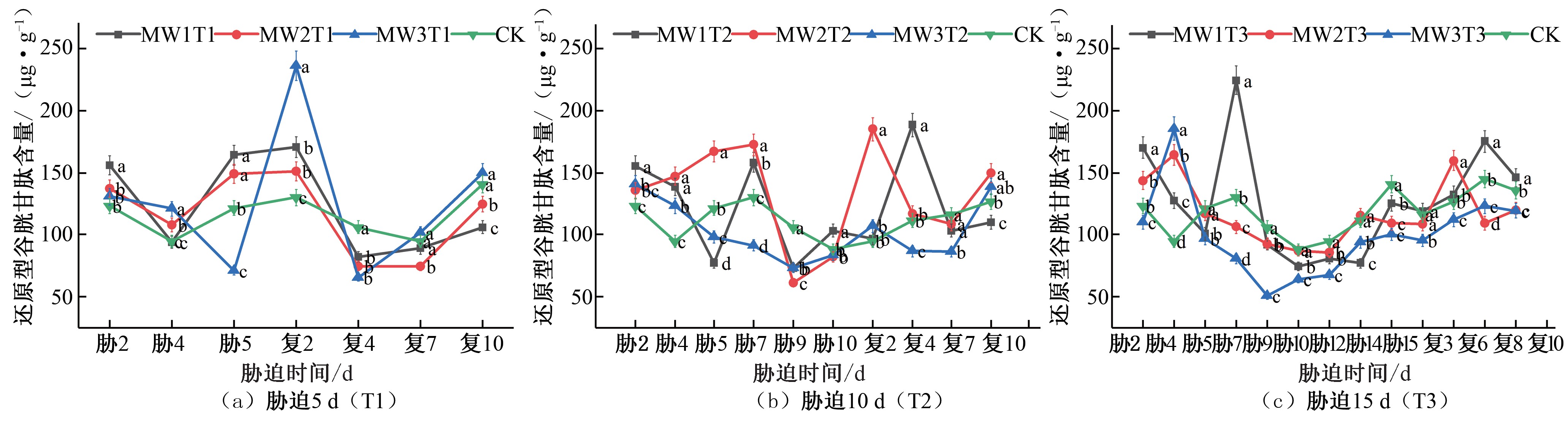
图9 夏玉米苗期还原型谷胱甘肽(GSH)含量于干旱胁迫-复水处理下的变化
Fig.9 Changes of GSH content in summer maize seedlings under drought stress and rehydration treatment
在夏玉米拔节期,胁迫时长为5 d 的处理GSH 含量均呈增加趋势,在胁迫末期达到峰值,BW1T1 和BW2T1 分别为CK 对照组的 1.22 倍和 0.96 倍,复水后逐渐降低,未出现补偿效应[见图10(a)]。胁迫时长为10 d 的处理胁迫期间的GSH 含量呈先增加后减少的趋势,BW1T2 的GSH 含量于胁迫第 3 天达到峰值,是CK 对照组的 1.23 倍,BW2T2 的GSH 含量未超过CK
对照组;复水后2 组处理GSH 含量均逐渐增加,但未出现补偿效应[见图10(b)]。胁迫时长为15 d 的处理,GSH 含量先增加后减少, BW1T3 的 GSH 含量始终低于 CK 对照组, BW2T3 的GSH 含量于胁迫第5 天达到峰值,是CK 对照组的1.06 倍,复水后2 组处理均始终低于CK 对照水平[见图10(c)]。
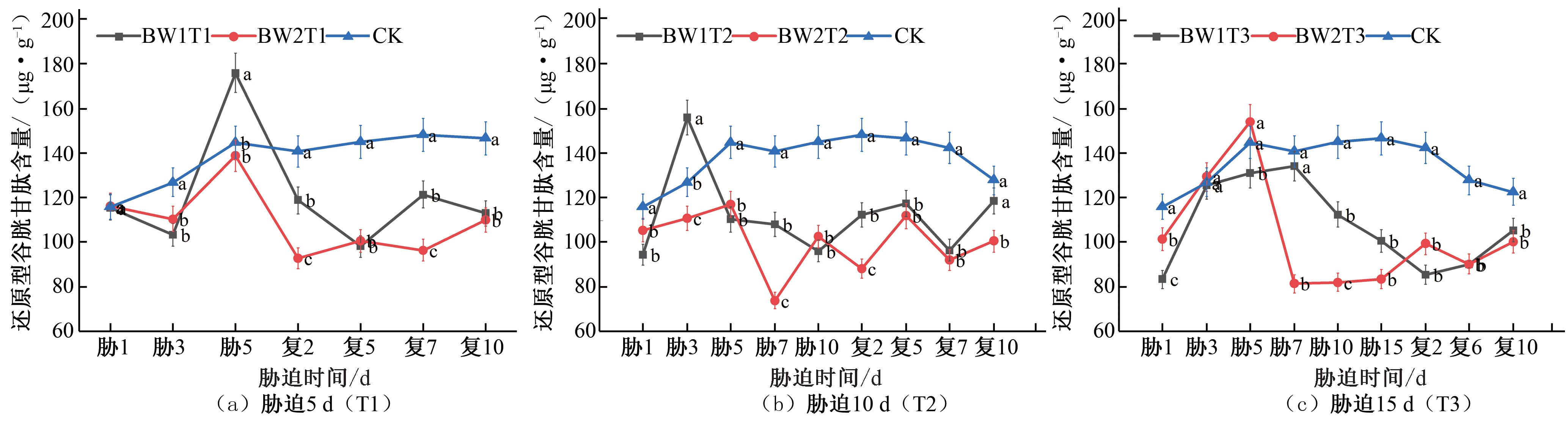
\删如餐#卷≈副隧斟
图10 夏玉米拔节期还原型谷胱甘肽(GSH)含量于干旱胁迫-复水处理下的变化
Fig.10 Changes of GSH cont\删如餐e#卷≈副隧n斟t in summer maize at jointing stage under drought stress\删如a餐#卷≈副隧n斟d rehydration treatment
综合对比夏玉米苗期和拔节期在干旱胁迫-复水过程中GSH 含量的变化响应,胁迫处理期间,在胁迫时长为 5 d 时, 轻度胁迫处理的GSH 含量高于中度和重度胁迫处理,胁迫时长 10 d 和 15 d 的处理组,中度胁迫的GSH 含量则是最高的, 植物体内GSH 含量随干旱胁迫程度加重而减小,这与王娟的研究结论一致[30]。复水后苗期的重度胁迫处理在胁迫 10 d 和
15 d 的处理组GSH 含量小于CK 对照处理,说明随着胁迫时长
3 结 论
综合比较夏玉米苗期和拔节期的MDA、SOD、POD、CAT 与GSH 的变化趋势,SOD、POD、CAT、GSH 在夏玉米的苗期均呈现出先增大后减小的趋势,在胁迫第4 天左右达到峰值。在拔节期,夏玉米的SOD、POD、CAT、GSH 呈现先增大后减小的趋势,均在胁迫 5~10 d 达到峰值。拔节期的各指标峰值
和胁迫程度的胁迫增时大间,/作d 物胞内代谢过程也受到了一定程度的 胁迫时的间活/性d(含量)均高于对应的苗期峰值。由此表明,胁夏迫玉时米间拔/节d
破坏。
期各个生化指标的调节能力均强于苗期,因此拔节期抗旱性
![]()
![]()
 要优于苗期。肖钢在对夏玉米的研究中也得出了类似结论[17]。 夏玉米的MDA 在胁迫期间均随胁迫时长和胁迫程度的增加不
要优于苗期。肖钢在对夏玉米的研究中也得出了类似结论[17]。 夏玉米的MDA 在胁迫期间均随胁迫时长和胁迫程度的增加不
: 断积累,在胁迫末期达到峰值,复水后随着胁迫缓解出现一
![]() 定补偿效应,最终逐渐恢复至CK 对照水平。通过MDA 含量的变化能初步判断细胞膜过氧化的程度和夏玉米对逆境条件的 反应强弱,也可以验证玉米抗氧化保护酶系统的响应机制。
定补偿效应,最终逐渐恢复至CK 对照水平。通过MDA 含量的变化能初步判断细胞膜过氧化的程度和夏玉米对逆境条件的 反应强弱,也可以验证玉米抗氧化保护酶系统的响应机制。
通过探究夏玉米生化指标对干旱-复水的响应变化,表明在对夏玉米进行节水灌溉时,从夏玉米各生化指标变化响应 来看,夏玉米苗期的抗旱能力不强,不宜开展持续时间较久
: 或干旱程度较重的亏水灌溉方案;夏玉米拔节期的抗旱能力 强于苗期,可以采用持续10 d 灌水量为55%~65% 土壤田间持
![]() 水量的胁迫处理方案进行灌溉,使得夏玉米受到的胁迫伤害 在作物自身可调节范围内,继而复水后出现补偿效应,以期 对夏玉米的高效节水灌溉提供理论支持。
水量的胁迫处理方案进行灌溉,使得夏玉米受到的胁迫伤害 在作物自身可调节范围内,继而复水后出现补偿效应,以期 对夏玉米的高效节水灌溉提供理论支持。
参考文献:
[ 1 ] 吴景社. 世界水土资源的潜力与粮食人口问题[J]. 世界农业,
: 1994(2):6-8.
[ 2 ] 王子豪,崔希东,郄志红,等 . 不同灌水处理对盆栽饲用谷子生长、耗水及产量的影响[J]. 中国农村水利水电,2019(4):69-73.
![]() [ 3 ] 苏治军,郝转芳,谢传晓,等 . 玉米dbf1 基因与耐旱相关性状的关联分析[J]. 植物遗传资源学报,2010,11(4):474-478.
[ 3 ] 苏治军,郝转芳,谢传晓,等 . 玉米dbf1 基因与耐旱相关性状的关联分析[J]. 植物遗传资源学报,2010,11(4):474-478.
[ 4 ] DAVID S G T,CHASCA T,HENNY O,et al. Adaptation to climate
 change and variability:farmer responses to intra-seasonal precipitation trends in South Africa[J]. Climatic Change,2007,83(3):301-322.
change and variability:farmer responses to intra-seasonal precipitation trends in South Africa[J]. Climatic Change,2007,83(3):301-322.
plant responses to drought - from genes to the whole plant. [J]. Functional plant biology :FPB,2003,30(3):2 365-2 384.
[ 12 ] 唐 建,冯 娟,刘云飞. 作物干旱复水补偿效应研究[J]. 湖北 :
农业科学,2021,60(1):10-13.
![]() [ 13 ] OLSEN L F,ISSINGER O,GUERRA B. The Yin and Yang of redox regulation[J]. Redox Report,2013,18(6):245-552.
[ 13 ] OLSEN L F,ISSINGER O,GUERRA B. The Yin and Yang of redox regulation[J]. Redox Report,2013,18(6):245-552.
[ 14 ] 胡国霞,马莲菊,陈 强,等. 植物抗氧化系统对水分胁迫及复水响应研究进展[J]. 安徽农业科学,2011,39(3):1 278-1 280.
[ 15 ] 刘婷婷. 玉米苗期干旱复水过程中叶片膜脂响应与干旱适应能力的关系[D]. 北京:中国科学院大学 中国科学院教育部水土保
持与生态环境研究中心,2018. :
![]() [ 16 ] WANG Y X,SUO B ,ZHAO P F,et al. Effect of exogenous abscisic acid on psbA expression at grain filling stage in two wheat cultivars under drought stress[J]. Acta Agronomica Sinica,2011,37 (8):1 372-1 377.
[ 16 ] WANG Y X,SUO B ,ZHAO P F,et al. Effect of exogenous abscisic acid on psbA expression at grain filling stage in two wheat cultivars under drought stress[J]. Acta Agronomica Sinica,2011,37 (8):1 372-1 377.
[ 17 ] 肖 钢. 不同时长干旱-复水对夏玉米生理调节机制及产量的影响[D]. 陕西杨凌:西北农林科技大学,2020.
[ 18 ] 邹原东,韩振芹,李志强. 水分胁迫对玉米苗期生理指标的影响
[J]. 安徽农业科学,2019,47(24):25-27. :
[ 19 ] 郭效龙,宋希云,裴玉贺,等. 玉米自交系萌发期和苗期抗旱性
指标的筛选[J]. 植物生理学报,2018,54(11):1 719-1 726.
![]() [ 20 ] 宫 亮,孙文涛,隽英华,等. 补充灌溉对玉米生理指标及水分利用效率的影响[J]. 节水灌溉,2017(1):9-11.
[ 20 ] 宫 亮,孙文涛,隽英华,等. 补充灌溉对玉米生理指标及水分利用效率的影响[J]. 节水灌溉,2017(1):9-11.
[ 21 ] 黄升谋. 干旱对植物的伤害及植物的抗旱机制[J]. 安徽农业科学,2009,37(22):10 370-10 372.
[ 22 ] 庄克章,吴荣华,张春艳,等. 苗期干旱及复水对玉米生长和生理
: [ 5 ] 于小兵,卢逸群,吉中会,等. 近45 a 来我国农业气象灾害变化特征及其对粮食产量的影响[J]. 长江流域资源与环境,2017,26(10): 1 700-1 710.
![]() [ 6 ] LU H D,XUE J Q,GUO D W. Efficacy of planting date adjustment as a cultivation strategy to cope with drought stress and increase rainfed maize yield and water-use efficiency[J]. Agricultural Water Management,2017,179(1):227-235.
[ 6 ] LU H D,XUE J Q,GUO D W. Efficacy of planting date adjustment as a cultivation strategy to cope with drought stress and increase rainfed maize yield and water-use efficiency[J]. Agricultural Water Management,2017,179(1):227-235.
[ 7 ] ZIYOMO C,BERNARDO R. Drought Tolerance in Maize:Indirect
: Selection through Secondary Traits versus Genomewide Selection[J].
Crop Science,2013,53(4):1 269-1 275.
[ 8 ] 刘 佳. 不同抗旱性玉米品种响应干旱及复水的光合生理机制
![]() [D]. 陕西杨凌:西北农林科技大学,2019.
[D]. 陕西杨凌:西北农林科技大学,2019.
[ 9 ] 刘 佳,杨永红,郝引川,等 . 干旱胁迫和复水处理对玉米叶片光合电子传递特性的影响[J]. 植物生理学报,2017,53(10): 1 877-1 884.
: [ 10 ] DAVID C N,MERLE F V,JOSEPH G B. The variable response of dryland corn yield to soil water content at planting[J]. Agricultural
Water Management,2008,96(2):330-336.
[ 11 ] CHAVES M M,MAROCO J O P,PEREIRA J O S. Understanding
 +
+
特性的影响[J]. 山东农业科学,2020,52(10):56-61. :
[ 23 ] 王伟玲,王 展,王晶英. 植物过氧化物酶活性测定方法优化[J]. 实验室研究与探索,2010,29(4):21-23.
![]() [ 24 ] 黄旭光,黄玲璞,王卫南,等. 干旱胁迫及复水对朱槿幼苗生理特性的影响[J]. 安徽农业科学,2020,48(21):137-139.
[ 24 ] 黄旭光,黄玲璞,王卫南,等. 干旱胁迫及复水对朱槿幼苗生理特性的影响[J]. 安徽农业科学,2020,48(21):137-139.
[ 25 ] VLADIMIR V B,MEI M W,SVETLANA V A,et al. Crystal
structure of manganese catalase from lactobacillus plantarum[J].
Structure,2001,9(8):725-738. :
[ 26 ] 李 冰,王文宗,林鸿佳,等. 超声波对过氧化氢酶酶活影响的
机理[J]. 华南理工大学学报( 自然科学版),2010,38(12):
129-134.
![]() [ 27 ] 刘 承,李佐同,杨克军,等. 水分胁迫及复水对不同耐旱性玉米生理特性的影响[J]. 植物生理学报,2015,51(5):702-708.
[ 27 ] 刘 承,李佐同,杨克军,等. 水分胁迫及复水对不同耐旱性玉米生理特性的影响[J]. 植物生理学报,2015,51(5):702-708.
[ 28 ] 樊跃平,于健春,余 跃,等. 谷胱甘肽的生理意义及其各种测定方法比较、评价[J]. 中国临床营养杂志,2003(2):58-61.
[ 29 ] 王玮玮,唐 亮,周文龙,等. 谷胱甘肽生物合成及代谢相关酶 :
的研究进展[J]. 中国生物工程杂志,2014,34(7):89-95.
[ 30 ] 王 娟,李德全,谷令坤. 不同抗旱性玉米幼苗根系抗氧化系统对水分胁迫的反应[J]. 西北植物学报,2002(2):77-82.
+
: :
k..+..+..+。+。+..+..+..+..+..+..+..+..+..+。+。+..+..+。+。+..+..+..+..+..+..+。+..+..+。+。+..+..+..+..+..+..+..+..+..+。+。+..+..+。+..+..+..√